

Abstract
Today, one of the biggest challenges in antibiotic research is a targeted prioritization of natural compound producer strains and an efficient dereplication process to avoid undesired rediscovery of already known substances. Thereby, genome sequence-driven mining strategies are often superior to wet-lab experiments because they are generally faster and less resource-intensive. In the current study, we report on the development of a novel in silico screening approach to evaluate the genetic potential of bacterial strains to produce protein synthesis inhibitors (PSI), which was termed the protein synthesis inhibitor ('psi’) target gene footprinting approach = Ψ-footprinting. The strategy is based on the occurrence of protein synthesis associated self-resistance genes in genome sequences of natural compound producers. The screening approach was applied to 406 genome sequences of actinomycetes strains from the DSMZ strain collection, resulting in the prioritization of 15 potential PSI producer strains. For twelve of them, extract samples showed protein synthesis inhibitory properties in in vitro transcription/translation assays. For four strains, namely Saccharopolyspora flava DSM 44771, Micromonospora aurantiaca DSM 43813, Nocardioides albertanoniae DSM 25218, and Geodermatophilus nigrescens DSM 45408, the protein synthesis inhibitory substance amicoumacin was identified by HPLC-MS analysis, which proved the functionality of the in silico screening approach.
Graphical Abstract
Graphical Abstract.
Ψ-footprinting approach.
INTRODUCTION
According to the World Health Organization (WHO), multidrug resistance is one of the major threats to public health (1). While the number of infections caused by multidrug-resistant bacteria is constantly increasing, there has been a steady decline in the discovery and development of new druggable anti-infectives over the past years (2,3). Thus, there is an urgent need for novel antibiotic active substances. One of the major challenges in current antibiotic screening programs is the fast dereplication of already known compounds. Indeed, most antibiotics were discovered during the golden age of antibiotic discovery from the 1940s to the 1960s (4). Back then, mainly the classical screening approach was applied, which included the isolation of soil-derived microorganisms that were screened for antimicrobial activity, a procedure known as the Waksman platform. However, many years of repeated application of this screening procedure mainly led to the rediscovery of already known substances, making it necessary to develop and apply new strategies for drug discovery. One such new approach, which we also refer to in the current manuscript, is represented by the mode of action (MoA)-based Bacillus subtilis bioreporter panel, recently reported by Wex et al. (5), which allows for fast and efficient combined bioactivity and MoA screening of antibiotic producer strains. The bioreporter assay employs selected biomarkers indicative of interference with the main bacterial metabolic pathways and thereby enables to selectively follow up and prioritize metabolic activities of interest (5). This MoA-based bioreporter assay was used to screen strains from the Tübingen actinomycetes strain collection for novel natural compounds (5). Preceding this study, the Tübingen strain collection had been subjected to a bioactivity test using the newly developed 96-well microplate system, designed to rapidly screen actinomycetes strain collections in agar plug assays (6). From a set of 500 natural product producers, 270 strains showed antibiotic activity against B. subtilis and were tested against the bioreporter panel sensing different types of metabolic stresses. Thereby, the bioreporter B. subtilis PbmrC-lacZ selectively responded to ribosomal stalling as a metabolic stress signal (5). Overall, six strains from the Tübingen strain collection, namely Tü 2975, Tü 3180, Tü 6430, KNN 49.3e, Tü 2108, and A 4/2 induced the B. subtilis PbmrC-lacZ bioreporter and were shown to produce protein synthesis inhibitors (PSIs), whereby Tü 2975 produced pristinamycin, Tü 3180 griseoviridin/viridogrisein, Tü 6430 pactamycin, KNN 49.3e amicetin, Tü 2108 berninamycin C, and A 4/2 cycloheximide (5). Cycloheximide was probably found rather accidentally, as A 4/2 only caused an ambiguous signal in the bioreporter assay and cycloheximide is known to act as eukaryotic PSI exclusively. In general, the bioreporter panel proved to be a highly efficient and sensitive tool for rapid compound identification and target deconvolution and allowed to identify natural products from strains of the Tübingen strains collection that had not been assigned to them before.
Other discovery efforts focus e.g. on data mining strategies involving genetic and analytical tools to exploit the so far hidden biosynthetic potential of the microbial natural compound producers. The most recent advances and achievements with a special focus on genome-guided discovery efforts have just recently been reviewed by Scherlach and Hertweck (7).
Regarding the antibiotic production capacity, actinomycetes are recognized as the most important source since they are the origin of up to 70% of today's therapeutically applied antibiotic agents (8–10). Even if these microorganisms have been extensively exploited over the past decades, they are still valuable sources for novel natural compounds. Bioinformatic analysis of >170 000 bacterial genomes and several thousand metagenome sequences suggests that overall only 3% of the genomic potential for natural product biosynthesis has been discovered so far and that especially the genus Streptomyces, belonging to the heterogeneous group of the actinomycetes, has a huge untapped potential for the production of novel secondary metabolites (11). The antibiotic encoding genes are usually organized as biosynthetic gene clusters (BGCs) in the genome of the producer strains (12). On average, Streptomyces genomes harbor 40 BGC (13), whereby the majority (∼90%) of all clusters is suggested to be silent, implying that the respective antibiotics are not produced under standard lab conditions (14). Bioinformatic tools and databases, such as antiSMASH, enable the prediction of BGCs in genome sequences (15) and allow genome mining of natural compound producers. Producer strains can be prioritized bioinformatically, e.g. by screening genome sequences for potential self-resistance genes. Specialized tools and databases such as the ‘Antibiotic Resistant Target Seeker’ (ARTS) allow for an automated prediction for potential resistance genes (16). The scientific background behind this data-mining attempt is that self-resistance mechanisms of natural compound producers can provide information of the MoA of the produced bioactive compound and can help to identify the encoding BGCs. For example, the molecular target of the cystobactamids was identified by studying the self-resistance mechanism in the myxobacterial producer organism Cystobacter sp. (17). Here, the gene cysO encodes a putative protein belonging to a family of pentapeptide repeat protein with a proven resistance function against type II topoisomerase drugs. cysO is part of the cystobactamid BGC and the implied function as potential resistance factor against type II topoisomerase drugs paved the way to disclose the MoA of the new family of cystobactamid antibiotics as topoisomerase inhibitors (18). The cystobactamid example shows that the search for potential resistance genes can help predict the MoA of the associated antibiotic and identify its encoding BGC. A similar example is shown by the anticancer agent salinosporamide A from Salinispora tropica CNB-440, a compound identified as a proteasome inhibitor due to the presence of a redundant proteasome subunit gene as part of the salinosporamide BGC (19). Following up on this observation, 86 Salinispora genomes were browsed for the occurrence of duplicated housekeeping genes that co-localized with BGCs. Thereby, a thiotetronic acid antibiotic BGC was found to be co-localized with a putative fatty acid synthase (FAS) resistance gene, which disclosed the MoA of the thiotetronic acid compounds as FAS inhibitors (20). Accordingly, for the antibiotics novobiocin (gyrase B), platensin (FabB/F) and griselimycin (DnaN), it is known that target-duplicated resistance genes are co-clustered with their encoding BGCs. These examples demonstrate the rationale of target-directed genome mining strategies (20). ARTS is a computational tool that takes this observation into account and enables the computer-aided search for potential self-resistance genes in bacterial genome sequences (21). ARTS predicts putative resistance genes based on three criteria: (i) duplication of genes, (ii) localization within a BGC and (iii) evidence for a horizontal gene transfer event (HGT). A defined set of core (‘housekeeping’) genes is used as a reference to identify essential core genes, defines duplication thresholds and infers HGT events. Traceability of HGT was implemented as a search criterion since antibiotic resistance genes are often transferred via HGT amongst bacteria and transfer events can be deduced from differences in the codon usage or GC content of DNA sequences compared to the rest of the genome (16). Altogether, ARTS represents an innovative tool that allows for a target-directed genome mining approach.
Based on the above-mentioned knowledge on BGC identification, we aimed to establish a genome sequence-based strategy to prioritize natural compound producers with a special emphasis on producers of PSIs. We focused on PSIs since the bacterial ribosome is a hotspot for many successful antibiotics, involving important drugs from different compound classes, e.g. aminoglycosides, macrolides, tetracyclines, lincosamides, pleuromutilins thiopeptides, or streptogramins (22). The bacterial ribosome is one of the most conserved functional units of the bacterial cell. It consists of the small 30S and the large 50S subunit, whereby the subunits are made up of >50 ribosomal proteins (r-proteins) and three rRNAs (23S, 16S and 5S). Ribosomal RNA represents the main target for antibiotics (23). A prominent example of a PSI is the aminoglycoside antibiotic streptomycin, which inhibits protein synthesis by interacting with the 16S rRNA and 20 ribosomal proteins of the 30S subunit (24). Streptomycin irreversibly binds to the 16S rRNA and the S12 protein, which are the main target sites (25). It inhibits the initiation process of protein synthesis (26) and causes decoding errors, which result in incorrect amino acids being incorporated into the nascent polypeptide chain (27). An example of a 50S subunit-binding PSI is the oxazolidinone antibiotic linezolid, which inhibits protein synthesis by binding to the peptidyl transferase center (PTC) of the ribosomal 50S subunit. Linezolid binds to the A-site of the PTC, interacting with various 23S rRNA nucleotides and the ribosomal S3 protein (28). Although the bacterial ribosome is a target for numerous different antibiotics, not all promising binding sites are therapeutically exploited so far, as exemplified by the PSIs evernimicin and negamycin. Negamycin is a small pseudopeptide with potent bioactivity against Gram-negative bacteria, which binds at a position near the A-site of the 30S subunit of the bacterial ribosome next to the binding site of the well-known PSI tetracycline (29). Evernimicin is an oligosaccharide antibiotic, which interacts with a specific set of nucleotides of the 23S rRNA of the 50S subunit, which are distinct from the binding sites of other known ribosome-targeting substances (30).
To search for novel natural compounds, which bind to the ribosome as a target, we have developed an in silico approach that allows to mine genomes for the occurrence and accumulation of protein synthesis associated-self-resistance genes. This method is called 'psi' protein synthesis inhibitor target gene footprinting = Ψ-footprinting. The method was validated by analyzing numerous genome sequences of known and recently identified PSI producers and then challenged with over 400 actinomycetes genomes from the DSMZ strain collection. From these, 15 strains were selected and probed for the production of natural compounds by HPLC-MS and protein synthesis inhibiting bioactivities by in vitro transcription/translation assays. For 12 of the 15 strains, protein synthesis inhibiting bioactivity was detected and for four of them the PSI was identified, proving the predictive power of the Ψ-footprinting approach.
MATERIALS AND METHODS
Bacterial strains, plasmids and cultivation conditions
The bacterial strains and plasmids used in this study are listed in Table 1. Escherichia coli NovaBlue (Novagen) was used as a cloning strain for the plasmid pET28-eGFP and E. coli BL21(DE3)pLysS (Novagen) was used for protein overexpression. For in vitro transcription/translation assays, E. coli strains were cultivated as reported previously (31). For cultivation and isolation of genomic DNA, actinomycetes strains were cultivated in 50 ml of R5 medium (32) in 500-ml Erlenmeyer flasks (with steel springs) on an orbital shaker (180 rpm) at 28°C.
Table 1.
Bacterial strains and plasmids
| Bacterial strain | Description | Source or reference |
|---|---|---|
| Escherichia coli | ||
| E. coli NovaBlue | endA1, hsdR17 (rK12– mK12 +), supE44, thi-1, recA1, gyrA96, relA1, lac, F' [proAB, lacIq, lacZΔM15, Tn10]; TetR | Novagen |
| E. coli BL21(DE3)/pLysS | F–ompT hsdSB(rB– mB–) gal dcm (DE3) pLysS (CmR) | Novagen |
| Bacillus subtilis | ||
| B. subtilis ATCC 6051 | Wild-type | American Type Culture Collection (ATCC) |
| B. subtilis 1S34 | amyE::pHJS105-PbmrC-lacZ | (5) |
| Actinomycetes strains | ||
| Streptomyces coelicolor M1146 | Δact Δred Δcpk Δcda | (53) |
| Streptomyces sp. Tü 2108 | Wild-type | Tübingen strain collection |
| Streptomyces sp. Tü 2975 | Wild-type | Tübingen strain collection |
| Streptomyces sp. Tü 3108 | Wild-type | Tübingen strain collection |
| Streptomyces sp. Tü 6430 | Wild-type | Tübingen strain collection |
| Streptomyces sp. A 4/2 | Wild-type | Tübingen strain collection |
| Streptomyces sp. KNN 49.3e | Wild-type | Tübingen strain collection |
| Nocardioides albertanoniae DSM 25218 | Wild-type | DSMZ strain collection |
| Micromonospora aurantiaca DSM 43813 | Wild-type | DSMZ strain collection |
| Micromonospora purpureochromogenes DSM 43821 | Wild-type | DSMZ strain collection |
| Actinomadura atramentaria DSM 43919 | Wild-type | DSMZ strain collection |
| Lentzea albidocapillata DSM 44073 | Wild-type | DSMZ strain collection |
| Rhodococcus koreensis DSM 44498 | Wild-type | DSMZ strain collection |
| Rhodococcus pyridinivorans DSM 44555 | Wild-type | DSMZ strain collection |
| Saccharopolyspora flava DSM 44771 | Wild-type | DSMZ strain collection |
| Rhodococcus kroppenstedtii DSM 44908 | Wild-type | DSMZ strain collection |
| Williamsia marianensis DSM 44944 | Wild-type | DSMZ strain collection |
| Jiangella alkaliphila DSM 45079 | Wild-type | DSMZ strain collection |
| Hoyosella altamirensis DSM 45258 | Wild-type | DSMZ strain collection |
| Geodermatophilus nigrescens DSM 45408 | Wild-type | DSMZ strain collection |
| Actinokineospora cianjurensis DSM 45657 | Wild-type | DSMZ strain collection |
| Micromonospora violae DSM 45888 | Wild-type | DSMZ strain collection |
| Plasmid | ||
| pET28a-eGFP | reporter plasmid (pET28a derivative) for in vitro transcription/translation assay; KanR | (31) |
For antibiotic production analyses, strains were cultivated in 50 ml inoculum medium (R5) at 28°C in 500 ml Erlenmeyer flasks (with steel springs) on an orbital shaker (180 rpm). After three days of cultivation, 5, 10 or 50 ml preculture was inoculated into 50, 100 or 200 ml of production medium (Supplementary Table S1), respectively. Cultures were grown for 4, 7 or 10 days at 28°C in 500 ml or 1 l Erlenmeyer flasks (with steel springs) on an orbital shaker (180 rpm).
Bacillus subtilis bioassay
Antibiotic activity was analyzed in agar plug diffusion assays using Bacillus subtilis ATCC 6051 as a test organism. Agar plugs of well-grown cultures of actinomycetes strains were placed on a B. subtilis test plate. In total, 5 μl of apramycin (50 μg/ml; Sigma) was applied as a positive control on filter discs. Plates were incubated overnight at 37°C and antibiotic activity was determined by measuring the diameter of the inhibition zone around the agar plugs and filter discs, respectively.
Target-specific bioreporter assay
Impairment of protein synthesis, more specifically the occurrence of translation arrest, was determined using the bioreporter strain B. subtilis 1S34 PbmrC-lacZ and agar-plug reporter assays were carried out as reported previously (5). Briefly, actinomycetes strains were cultivated for 7–10 days at 28°C on the indicated agar plates. Then, grown agar plugs of the well-grown strains were transferred to an empty petri dish before embedding them in B. subtilis 1S34 PbmrC-lacZ-containing soft agar. The agar was supplemented 150 μg/ml X-Gal (Thermo Scientific), that gives blue colorization in plates after promoter induction. Extracts or purified compounds were directly spotted on solidified soft agar plates containing B. subtilis 1S34 PbmrC-lacZ. Antibiotic discs containing 10 μg of chloramphenicol (Oxoid) were used as a positive control. Agar plates were analyzed for promoter induction after overnight incubation at 37°C. PbmrC induction was indicated by a blue halo surrounding the zone of inhibition of the respective test sample.
Genomic DNA isolation and sequencing
Isolation of genomic DNA and genome sequencing of the strains Tü 2975, Tü 3180, Tü 6340 and KNN 49.3e was performed as described in Hennrich et al. (33) and for strains Tü 2108 and A 4/2 as described in Zecher et al. (34). Genome sequence-specific information is listed in Supplementary Table S2.
Type Strain Genome Server (TYGS)
The phylogenomic analysis of selected strains was carried out with the Type (Strain) Genome Server (TYGS) v. 1.0, a free bioinformatics tool for whole-genome-based taxonomic analysis (35). Phylogenetic classification using TYGS is based on a genome database that contains the genomic, taxonomic, and nomenclatural data of all currently available type strains. The database is constantly updated (https://tygs.dsmz.de/). The TYGS platform allows phylogenetic analyses based on the full-length genome sequence of the strain of interest, which is compared with a database of type strain genomes. Thereby, TYGS provides information on the similarity of the strain to its nearest related type strain with the help of digital DNA:DNA hybridization (dDDH) values calculated by the Genome-to-Genome Distance Calculator (GGDC) 2.1 (http://ggdc.dsmz.de) (36) and determines differences in genomic G + C contents.
ARTS analysis
The Antibiotic Resistance Target Seeker (ARTS) web tool v. 2.0 (21) was used to screen for potential self-resistance genes in genome sequences of natural compound producers. Genome sequences or accession numbers were submitted to the ARTS web-tool. The reference set was selected for Actinobacteria and the exploration mode was used. For all analyses, default settings were used.
AntiSMASH analysis
The antiSMASH web tool v. 6.0 (15) was used to analyze whole genome sequences for the presence of BGCs. Gene cluster similarity is displayed in % and indicates the number of genes similar to a known cluster. Genes are similar if a BLAST alignment yields an e-value <1 × 10–5 and if sequence identity is >30%. In addition, the shortest alignment must encompass >25% of the sequence. If all genes of a known cluster can be found in the query cluster, the sequence similarity is 100%. Similarity decreases if fewer genes of the known cluster can be found in the query cluster (37). Default settings were used for all analyses.
In vitro transcription/translation (ivTT) assay
The in vitro transcription/translation (ivTT) assay was performed as reported previously (31). Briefly, an S12 extract was prepared from E. coli BL21(DE3)pLysS and mixed with nucleotides and amino acids as substrates for the transcription and translation process. Fructose-1,6-bisphosphate was added as an energy supply and pET28-egfp as reporter plasmid. In total, 85 μl of 2.5× ivTT buffer, 60 μl of the S12 extract, 20 μl of an amino acid mixture (2 mM each of all 20 proteinogenic amino acids), and 20 μl of an energy mixture (1.2 mM AMP, 0.85 mM each of CMP, GMP, and UMP, 0.5 mM IPTG, 34 mg/ml l-5-formyl-THF and 33 mM fructose-1,6- bisphosphate) served as the master mix for one sample of the ivTT assay (31). To test the actinomycetes culture extracts for translation inhibition, 30 μl of methanolic extracts were added to each well of a 96-well microtiter plate and incubated for 30 min at 37°C to evaporate the methanolic solvent. The master mix and 10 μl of the reporter plasmid pET28-egfp (300 ng/μl) were added to each well to a final volume of 100 μl, thereby starting the reaction. All measurements were performed in triplicate using the same preparation of E. coli S12 extract. As positive controls, 5 μl of tetracycline (15 mg/ml; Sigma) or 5 μl of apramycin (50 mg/ml; Sigma) were used. The assay was incubated for approximately 2 h at 37°C. Fluorescence deriving from eGFP expression was measured at 485/520 nm excitation/emission with a Tecan Infinite M200 Pro device and analyzed with the i-control™ Microplate Reader Software 1.11 (Tecan).
Compound extraction from actinomycetes cultures
After cultivation of actinomycetes strains for 1–10 days in production medium, 5 or 50 ml of culture were extracted with 5, or 50 ml ethyl acetate, respectively, for 30 min at room temperature. Ethyl acetate samples were evaporated in vacuo completely and then redissolved in 0.25 or 1.25 ml of methanol, respectively. Methanolic extracts were used for bioassays and high-performance liquid chromatography/mass spectrometry (HPLC–MS) analysis, were stored at 4°C or frozen at −20°C for later analysis. As controls, pure media extracts were prepared at a ratio of 1:1 media/ethyl acetate.
Semi-preparative purification by HPLC
For compound isolation, semi-preparative HPLC was carried out with an Agilent Technologies 1260 Infinity (Agilent Technologies, Waldbronn, Germany). A sample volume of 100 μl was injected at a flow rate of 1.5 ml/min on a Luna C18 column (5 μm, 100 Å, 4.6 mm × 250 mm) and a linear gradient from 5% solvent A (0.1% formic acid in water) to 100% solvent B (0.1% formic acid in acetonitrile) over 25 min at 25°C. 1.5 ml volume of each fraction was collected time based every minute.
HPLC–MS analysis
HPLC analyses were performed as described previously with a HP1090M system and ChemStation 3D software rev. A.08.03 (Agilent Technologies, Waldbronn, Germany) on a Nucleosil C18 column (5 mm, 125 mm × 3 mm) fitted with a precolumn (20 × 3 mm) and with a flow rate of 850 μl/min (38). HPLC–MS was performed with an Agilent 1200 series chromatography system (binary pump, high performance autosampler, DAD-detector) coupled to a LC/MSD Ultra Trap System XCT 6330 (Agilent Technologies, Waldbronn, Germany) (38). Samples were injected onto a Nucleosil 100 C18 column (3 μm, 100 × 2 mm) fitted with a precolumn (3 μm, 10 × 2 mm) at a flow rate of 400 μl/min and a linear gradient from 10% solvent A (0.1% formic acid in water) to 100% solvent B (0.06% formic acid in acetonitrile) over 15 min at 40°C.
For the dereplication process, the UV-DAD spectra of the extracts/fractions were analyzed first for relevant signals. For this purpose, the peaks in the HPLC chromatogram were individually analyzed by comparing the chromatogram with an in-house HPLC–UV–Vis database (39). Peaks showing a coincident UV–Vis entry with a possible PSI substance were followed up. The masses of the possible PSI compounds were compared with the Dictionary of Natural Products as an external database (40). A substance was classified as ‘clearly identified’, if the match with the internal database and the external database confirmed the UV–Vis and MS criteria.
RESULTS AND DISCUSSION
Genome mining of strain Tü 2108 leads to the identification of a berninamycin BGC and provides information on berninamycin derivative biosynthesis
In a previous study, we established a bioreporter assay panel, suitable for MoA profiling of antibacterial agents, which are secreted into the agar by producer strains. This procedure was used to screen strains from the Tübingen actinomycetes strain collection for novel natural compounds (5). Here, we used the bioreporter strain B. subtilis PbmrC-lacZ that specifically responds to protein synthesis inhibition, or more precisely to ribosomal stalling, as a metabolic stress signal (5). Antibiotic activity was indicated by a zone of growth inhibition in the bioreporter lawn, while the inhibitory effect on protein synthesis was signaled by a blue halo around the inhibition zone as a result of brmC promoter activation that drives β-galactosidase reporter gene expression and associated color change (5).
Strain Tü 2108 emerged in this MoA-informed screening process as one of the strains, which yielded a large inhibition zone and a clear blue halo (Figure 1). The implicated PSI produced by the strain was identified as berninamycin C by HPLC-MS analysis (5). For the characterization of the Tü 2108 producer organism, the genomic DNA was sequenced and the full-length genome sequence was analyzed bioinformatically for phylogenetic classification and identification of BGCs. Genome-sequence-based phylogenetic analysis was performed using the Type (Strain) Genome Server v. 1.0 (TYGS) (https://tygs.dsmz.de) (35). TYGS analysis with the genome sequence of Tü 2108 revealed that the strain is most closely related to the type strain Streptomyces atroolivaceus NRRL ISP-5137T (Figure 2). The dDDH value (formula d4) between Tü 2108 and S. atroolivaceus NRRL ISP-5137T was 84.6%, which is above the threshold of 70% commonly used for species delineation. Thus, Tü 2108 belongs to the same species as S. atroolivaceus NRRL ISP-5137T, which has already been described as a berninamycin producer strain (41). To investigate if the berninamycin BGC is present in Tü 2108, the genome sequence was analyzed with the bioinformatic tool antiSMASH v. 6.0 (15), which led to the identification of 38 BGCs, whereby the predicted BGC located on region 6_3 (BGC 6_3) showed ∼100% similarity to the berninamycin A BGC of the berninamycin producer strain Streptomyces bernensis (Supplementary Figure S1, Figure 3). Manual sequence analysis revealed that the antiSMASH database contained an incomplete berninamycin A BGC as a reference (MIBiG accession BGC0001472), lacking the gene berH from the berninamycin A BGC (berA-J) as initially described by Malcolmson et al. (42).This was found after BLAST analysis of each individual gene of BGC 6_3 from Tü 2108 and comparisons with the respective BGC genes from S. bernensis (Table 2). Thus, all genes essential for berninamycin biosynthesis are present in Tü 2108.
Figure 1.
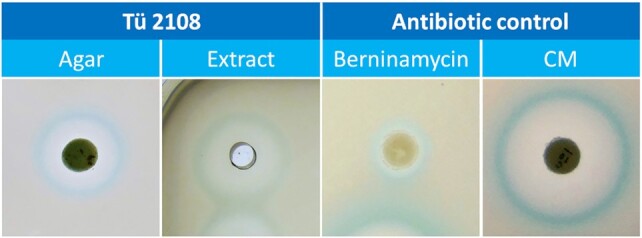
Agar-based reporter screening with samples from Tü 2108 and bioreporter test strain B. subtilis PbmrC-lacZ. 1. panel: Tü 2108-grown OM agar plug; 2. panel: 5 μl culture extract from Tü 2108 grown in R5 medium; 3.-4. panel: Pure antibiotics (50 μg berninamycin and 10 μg chloramphenicol (CM)) were prepared on filter discs as controls.
Figure 2.

Whole-genome sequence tree generated with the TYGS web server for strain Tü 2108 and closely related species. Tree inferred with FastME from GBDP distances calculated from genome sequences. The branch lengths are scaled in terms of GBDP distance formula d5. The numbers above branches are GBDP pseudo-bootstrap support values >60% from 100 replications, with an average branch support of 84.9%.
Figure 3.

Cluster comparison between S. bernensis berninamycin A gene cluster and Tü 2108 region 6_3. Black: berninamycin biosynthesis genes. Yellow: ribosomal genes.
Table 2.
Berninamycin-related genes and their described function. Tü 2108 region 6_3 ORFs is compared to the homologous S. bernensis genes. Gene identity (ID) and similarity (SM) of compared amino acid sequences are shown in %. ORF = open reading frame
| Tü 2108 region 6_3 ORFs | Predicted gene product | Homologous S. bernensis gene | ID/SM (%) | Reference |
|---|---|---|---|---|
| 01231 | LmbE family protein | acyl transferase | 100/100 | AGN11661 |
| 01232 | rRNA methyl transferase | berJ | 99/100 | AGN11674 |
| 01233 | NocA homolog (C-terminal amide) | berI | 99/99 | AGN11673 |
| 01234 | cytochrome P450 | berH | 99/99 | KC894738 |
| 01235 | lanthipeptide dehydrogenase | berC | 99/100 | AGN11672 |
| 01236 | lanthipeptide dehydrogenase | berB | 97/98 | AGN11671 |
| 01237 | berninamycin prepeptide gene | berA | 98/100 | AGN11670 |
| 01238 | pyridine-forming | berD | 93/96 | AGN11669 |
| 01239 | YcaO cyclodehydratase | berG2 | 99/99 | AGN11668 |
| 01240 | YcaO cyclodehydratase | berG1 | 99/99 | AGN11667 |
| 01241 | McbC dehydrogenase | berE2 | 98/99 | AGN11665 |
| 01242 | McbC dehydrogenase | berE1 | 99/98 | AGN11666 |
| 01243 | elongation factor Tu | tufA/B | 100/100 | AGN11675 |
| 01244 | elongation factor G | fusA | 99/100 | AGN11662 |
| 01245 | 30S ribosomal protein S7 | rpsG | 100/100 | AGN11664 |
| 01246 | 30S ribosomal protein S12 | rpsL | 99/100 | AGN11663 |
Bioinformatic analyses disclose that the berninamycin BGC is surrounded by potential self-resistance genes
Berninamycin belongs to the class of pyridine-containing thiopeptides and targets the 50S ribosomal subunit in a similar manner as described for thiostrepton (42). Several berninamycin producer strains are known, whereby the production of berninamycin derivatives differs in the various producer strains (Supplemental material Figure S3). A comparison between the known berninamycin BGC from S. bernensis and the BGC from Tü 2108 revealed that the predicted gene products show high amino acid sequence homologies (90–100%) amongst each other (Table 2). To identify potential self-resistance genes for strain Tü 2108, the genome sequence was analyzed with the bioinformatics software tool ARTS v. 2.0. As mentioned above, ARTS detects potential self-resistance genes based on the following individual three criteria: (i) duplicated core genes, (ii) core genes localized within a BGC and (iii) core genes with incongruent phylogeny (16). For Tü 2108, the suggested self-resistance genes were sorted with regard to genes associated with protein synthesis as a metabolic function. Concerning protein synthesis associated genes, ARTS employs a reference set of 100 core genes related to protein synthesis (16). For Tü 2108 it was found that 15 protein synthesis-associated genes met the above mentioned criteria for resistance patterns: In total, nine duplicated core genes, 13 core genes located in a BGC, and six phylogenetically distinct core genes were predicted (Table 3). The majority of the detected core genes present in a BGC, mapped to the berninamycin cluster BGC 6_3 (Table 2). Here, the core genes are organized at the left and right borders of the berninamycin BGC 6_3 (Figure 3) and code for potential ribosomal proteins (L4, S12, S10, uS7), as well as elongation factors G and TU (Table 2). For instance, ORF 01243 encoding the putative elongation factor TU (EF-TU) was duplicated and phylogenetically distinct from another putative EF-TU encoding gene (TIGR00485) present in the Tü 2108 genome. A similar genetic organization was found for the berninamycin A BGC (berA-J) in S. bernensis (Figure 3). Notably, also a berJ homologous gene is present within BGC 6_3 of Tü 2108, which codes for a potential 23S rRNA methyltransferase, an enzyme well-known for conferring resistance to ribosome-targeting drugs (43). However, this gene, which most likely represents the actual berninamycin self-resistance gene, was not detected with ARTS, which is due to the fact that it was not included in the list of core genes. The predominant occurrence of these protein synthesis-associated core genes in the proximity of the berninamycin BGC indicates a potential linkage to self-resistance mechanisms. This finding supports the hypothesis that resistance genes can provide an indication of the mode of action of a BGC-encoded secondary metabolite.
Table 3.
Protein synthesis core genes of Tü 2108 identified with ARTS. Following criteria are listed: gene duplication, phylogenetic gene difference, occurrence of core genes in a biosynthetic gene cluster (BCG). +: criterion fulfilled, –: criterion not fulfilled. For BGC prediction, only region 6_3 (berninamycin BGC) is listed
| Gene | Description | Duplication | Phylogeny | BGC | Accession ID |
|---|---|---|---|---|---|
| rpsR | 30S ribosomal protein S18 | + | + | + | TIGR00165 |
| yciO | tRNA threonylcarbamoyl adenosine modification protein, Sua5/YciO/YrdC/YwlC family | + | + | + | TIGR00057 |
| rpmF | 50S ribosomal protein L32 | + | + | + | TIGR01031 |
| rpmE | 50S ribosomal protein L31 | + | + | + | TIGR00105 |
| tufA/B | elongation factor Tu | + | + | 6_3 | TIGR00485 |
| rpmB | 50S ribosomal protein L28 | + | - | + | TIGR00009 |
| rpmG | 50S ribosomal protein L33 | + | - | + | TIGR01023 |
| rpsD | 50S ribosomal protein L4 | - | - | 6_3 | TIGR03953 |
| rpsL | 30S ribosomal protein S12 | - | - | 6_3 | TIGR00981 |
| rpsJ | 30S ribosomal protein S10 | - | - | 6_3 | TIGR01049 |
| rpsO | 30S ribosomal protein S15 | - | - | + | TIGR00952 |
| rpsG | 30S ribosomal protein uS7 | - | - | 6_3 | TIGR01029 |
| fusA | elongation factor G | - | - | 6_3 | TIGR00484 |
| rrsS | tryptophan-tRNA ligase | + | + | - | TIGR00233 |
| rsgA | ribosome small subunit-dependent GTPase A | + | - | - | TIGR00157 |
Development of a genome sequenced-based screening approach for the identification of PSI producer strains
Based on the knowledge gained above, an in silico screening approach was developed, capitalizing on the occurrence of protein synthesis associated self-resistance genes to evaluate the genetic potential of bacterial strains to produce a PSI. To validate the approach, genome sequences of 47 known PSI producers were analyzed with ARTS. Here, it was found that several known PSI producers showed high numbers (>20) of protein synthesis core genes, fulfilling at least one of the three criteria mentioned for the prediction of self-resistance genes, defined as ‘hit genes’. For example, the anisomycin producer Streptomyces roseochromogenus displayed 42 hit genes and the erythromycin producer Saccharopolyspora erythraea 51 hit genes (Table 4). The majority of the identified hit genes of the PSI producers were characterized as phylogenetic incongruencies (76%) and only a minor part represented core gene duplications (46%) or gene localization within a BGC (26%) (Supplementary Table S3). Among the hit genes, seven core genes appeared regularly, which were rpsE, rpsL, rplC, rplD, rplK, rplQ and rplV encoding for the putative ribosomal proteins S5, S12, L3, L4, L11, L17 and L22, respectively (Table 4). For all these genes, it was shown in previous work that they play an essential role in conferring resistance against PSIs. For example, mutations in rpsE, rpsL, and rplC resulted in altered S5, S12 and L3 proteins that conferred resistance against spectinomycin, streptomycin, and linezolid/tiamulin, respectively, in E. coli (44–46). Mutations in rplD and rplV were shown to result in altered L4 and L22 proteins, respectively, conferring resistance to erythromycin in E. coli (47,48). Mutations in rplK resulted in an altered L11 protein conferring resistance to thiostrepton in Thermus thermophilus (49) and mutations in rplQ resulted in an altered L17 protein conferring resistance to erythromycin in B. subtilis (50). According to these observations, the seven ribosomal genes were regarded as PSI 'resistance indicator genes’ (RI genes). The RI genes were found to be mainly present in PSI producer strains with a high number of PSI hit genes, e.g. S. roseochromogenus and S. erythraea with six and five RI genes, respectively (Table 4). However, some of the RI genes were also found in PSI producers with a lower number of hit genes, such as Streptomyces pristinaespiralis PR11 and Streptomyces pactum, with only one RI gene each. Furthermore, there were also examples of known PSI producers that showed neither a high abundance of hit genes nor any RI gene (Table 4). To examine whether the accumulation of hit and RI genes is a specific phenomenon for PSI producers, we additionally analyzed numerous genome sequences of antibiotic producers, which are known not to produce PSIs as a comparison. Thereby, it was found that the non-PSI producers had generally low numbers of protein synthesis-related hit genes (<20) and none of the strains contained any of the seven RI genes (Table 5). Thus, we concluded that if a bacterial strain has (i) a large number of PSI hit genes (>20) and in addition (ii) two or more RI gene(s) it is likely to be a PSI producer strain.
Table 4.
List of protein synthesis inhibitor (PSI) producer strains and their respective PSIs. Strains are ordered according to the occurrence of protein synthesis ‘hit genes’. Abundance of ‘resistance indicator’ (RI) genes/encoded proteins is highlighted as plus. Gene/protein absence is indicated as minus
| Strain | Antibiotic (PSI) | Hit genes | rpsE/S5 | rpsL/S12 | rplC/L3 | rplD/L4 | rplK/L11 | rplQ/L17 | rplV/L22 | RI genes | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Thermus thermophilus HB8 | Paromomycin | 75 | + | + | - | + | + | - | - | 4 | AP0082 |
| Saccharopolyspora erythraea NRRL 2338 | Erythromycin | 51 | + | + | - | + | + | + | - | 5 | NC_009142 |
| Micromonospora echinospora DSM 43816 | Gentamicin | 49 | - | + | - | - | - | + | + | 3 | NZ_LT60748 |
| Kitasatospora setae KM-6054 | Kirromycin | 48 | - | - | + | + | + | + | + | 5 | AP010968 |
| Sphaerisporangium cinnabarinum ATCC 31213 | Dityromycin | 47 | - | + | - | + | - | - | - | 2 | QAPD00000000 |
| Micromonospora inyonensis NRRL 3292 | Sisomicin | 46 | - | - | - | - | - | + | + | 2 | GCA_900091415 |
| Brevibacillus brevis X23 | Edeine | 43 | + | - | - | + | - | - | - | 2 | NZ_CP023474 |
| Streptomyces roseochromogenus | Anisomycin | 42 | + | + | + | + | - | + | + | 6 | NZ_CM0022 |
| Micromonospora carbonaceae | Evernimicin | 40 | + | - | - | - | - | - | + | 2 | NZ_CP058322 |
| Streptomyces cattleya DSM 46488 | Kirromycin | 28 | + | + | - | + | - | - | + | 4 | CP003219 |
| Streptomyces cattleya NRRL 8057 | Kirromycin | 27 | - | + | - | + | - | - | + | 3 | FQ859185 |
| Rhodococcus erythropolis PR4 | Chloramphenicol | 24 | - | - | - | + | - | - | + | 2 | AP008957 |
| Rhodococcus pyridinivorans SB3094 | Chalcomycin | 21 | - | + | - | + | - | - | + | 3 | CP006996 |
| Streptomyces griseochromogenes ATCC 14511 | Blasticidin S | 19 | - | - | - | - | - | - | - | - | NZ_CP0162 |
| Xenorhabdus nematophila SII | Odilorhabdins | 19 | + | - | - | - | + | - | - | 2 | NZ_CP060401 |
| Streptomyces kasugaensis AM-2504 | Dityromycin | 17 | - | - | - | - | - | - | - | - | SIXH01000000 |
| Streptomyces rimosus ATCC 10970 | Oxytetracyclin | 17 | - | + | - | - | - | - | - | 1 | NZ_CP0236 |
| Streptomyces bikiniensis NRRL B-1049 | Chalcomycin | 15 | - | - | - | - | - | - | - | - | JNWL00000000 |
| Streptomyces collinus Tü 365 | Kirromycin | 15 | - | - | - | - | - | - | - | - | CP006259.1 |
| Streptomyces lateritius JCM 4389 | Granaticin | 15 | - | - | - | - | - | - | - | - | BMTO00000000 |
| Kitasatospora aureofaciens DM-1 | Chlortetracycline | 14 | - | - | - | - | - | - | - | - | NZ_CP0205 |
| Streptomyces albulus NK660 | Anisomycin | 14 | - | - | - | - | - | - | - | - | NZ_CP0075 |
| Streptomyces pristinaespiralis PR11 | Pristinamycin | 14 | - | + | - | - | - | - | - | 1 | CP059696 |
| Streptomyces scabiei 87.22 | Bottromycin A2 | 14 | - | - | - | - | - | - | - | - | NC_013929 |
| Streptomyces albireticuli MDJK11 | Spiramycin | 13 | - | - | - | - | - | - | - | - | NZ_CP0217 |
| Streptomyces globosus LZH-48 | Factumycin | 13 | - | - | - | - | - | - | - | - | CP030862 |
| Streptomyces lincolnensis NRRL 2936 | Lincomycin | 13 | - | - | - | - | - | - | - | - | NZ_CP016438 |
| Streptomyces pactum ACT12 | Pactamycin | 13 | - | + | - | - | - | - | - | 1 | NZ_CP019724 |
| Streptomyces venezuelae ATCC 10712 | Chloramphenicol | 13 | - | - | - | - | - | - | - | - | NC_018750 |
| Streptomyces vietnamensis GIM4.0001 | Granaticin | 13 | - | - | - | - | - | - | - | - | NZ_CP0104 |
| Streptomyces viridochromogenes Tü 57 | Avilamycin | 13 | - | - | - | - | - | - | - | - | NZ_AMLP0000000 |
| Streptomyces xinghaiensis S187 | Neomycin | 13 | - | - | - | - | - | - | - | - | NZ_CP023202 |
| Streptomyces ambofaciens DSM 40697 | Midecamycin | 12 | - | - | - | - | - | - | - | - | NZ_CP012949 |
| Streptomyces chrestomyceticus TBRC 1925 | Paromomycin | 12 | - | - | - | - | - | - | - | - | JAEAGG010000010 |
| Streptomyces pristinaespiralis ATCC 25486 | Pristinamycin | 12 | - | - | - | - | - | - | - | - | CM000950 |
| Streptoverticillium mobaraenses NBRC 13819 | Pulvomycin | 12 | - | - | - | - | - | - | - | - | NZ_CP072827 |
| Streptomyces lavendulae CCM 3239 | Streptothricin | 11 | - | - | - | - | + | - | - | 1 | NZ_CP024985 |
| Streptomyces lincolnensis NRRL 2936 | Lincomycin | 11 | - | - | - | - | - | - | - | - | NZ_CP0164 |
| Streptomyces antibioticus ATCC11891 | Oleandomycin | 10 | - | - | - | - | - | - | - | - | NZ_CP050692 |
| Streptomyces bottropensis ATCC 25435 | Bottromycin A2 | 10 | - | - | - | - | - | - | - | - | NZ_ARTP0000000 |
| Streptomyces laurentii ATCC 31255 | Thiostrepton | 10 | - | - | - | - | - | - | - | - | AP017424 |
| Streptomyces vinaceus NRRL ISP-5257 | Viomycin | 10 | - | - | - | - | - | - | - | - | NRRL ISP-5257 |
| Streptomyces violaceoruber S21 | Viomycin | 10 | - | - | - | - | - | - | - | - | NZ_CP020570 |
| Streptomyces fradiae ATCC 10745 | Neomycin | 9 | - | - | - | - | - | - | - | - | NZ_CP023696 |
| Streptomyces griseus NBRC 13350 | Streptomycin | 8 | - | - | - | - | - | - | - | - | NC_010572 |
| Streptomyces sparsogenes ATCC 25498 | Sparsomycin | 8 | - | - | - | - | - | - | - | - | MAXF00000000 |
Table 5.
List of non-PSI producer strains and their known antibiotic products. Strains are ordered according to the occurrence of protein synthesis ‘hit genes’. Abundance of ‘resistance indicator’ genes/encoded proteins is highlighted as plus. Gene/protein absence is indicated black minus
| Strain | Antibiotic (non-PSI) | Hit genes | rpsE/S5 | rpsL/S12 | rplC/L3 | rplD/L4 | rplK/L11 | rplQ/L17 | rplV/L22 | RI genes | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Actinoplanes friuliensis DSM 7358 | Friulimicin | 17 | - | - | - | - | - | - | - | - | CP006272 |
| Amycolatopsis mediterranei S699 | Rifamycin | 17 | - | - | - | - | - | - | - | - | CP002896 |
| Mycobacterium avium 104 | Glycopeptidolipid, Mycobactin | 15 | - | - | - | - | - | - | - | - | CP000479 |
| Rhodococcus jostii RHA1 | Rhodochelin | 15 | - | - | - | - | - | - | - | - | CP000431 |
| Amycolatopsis orientalis HCCB 10007 | Vancomycin | 14 | - | - | - | - | - | - | - | - | CP003410 |
| Streptomyces brunneus CR22 | Feglimycin | 14 | - | - | - | - | - | - | - | - | NZ_CP034463 |
| Streptomyces violaceusniger Tü 4113 | Nigericin | 14 | - | - | - | - | - | - | - | - | CP002994 |
| Streptomyces chartreusis NRRL 3882 | Tunicamycin | 13 | - | - | - | - | - | - | - | - | NZ_LT963352 |
| Streptomyces parvulus 2297 | Actinomycin D Dactinomycin | 13 | - | - | - | - | - | - | - | - | NZ_CP015866 |
| Streptomyces viridochrom. DSM 40736 | Phosphinothricin | 13 | - | - | - | - | - | - | - | - | GG657757 |
| Streptomyces davawensis JCM 4913 | Roseoflavin | 12 | - | - | - | - | - | - | - | - | HE971709 |
| Streptomyces formicae KY5 | Formicamycins A-M | 12 | - | - | - | - | - | - | - | - | NZ_CP022685 |
| Streptomyces avermitilis MA-4680 | Avermectin Oligomycin | 11 | - | - | - | - | - | - | - | - | NC_003155 |
| Streptomyces bingchenggensis BCW-1 | Meridamycin | 11 | - | - | - | - | - | - | - | - | NC_016582 |
| Streptomyces coelicolor A3(2) | Actinorhodin Undecylprodigiosin | 11 | - | - | - | - | - | - | - | - | NC_003888 |
| Streptomyces lividans TK24 | Actinorhodin Undecylprodigiosin | 11 | - | - | - | - | - | - | - | - | NZ_GG657756 |
| Streptomyces malaysiensis DSM 4137 | Azalomycin F | 11 | - | - | - | - | - | - | - | - | NZ_CP023992 |
| Streptomyces niveus NCIMB 11891 | Novobiocin | 11 | - | - | - | - | - | - | - | - | NZ_CM002280 |
| Salinispora tropica CNB-440 | Salinisporamide A | 11 | - | - | - | - | - | - | - | - | CP000667 |
| Streptomyces albus DSM 41398 | Salinomycin | 10 | - | - | - | - | - | - | - | - | NZ_CP010519 |
| Streptomyces flavogriseus ATCC 33331 | Carbapenem | 10 | - | - | - | - | - | - | - | - | CP002475 |
| Streptomyces pratensis ATCC 33331 | Carbapenem | 10 | - | - | - | - | - | - | - | - | NC_016114 |
| Thermobifida fusca YX | Fuscachelin | 10 | - | - | - | - | - | - | - | - | CP000088 |
| Streptomyces clavuligerus ATCC 27064 | Tunicamycin | 9 | - | - | - | - | - | - | - | - | NZ_CP027858 |
| Streptomyces tsukubaensis NRRL 18488 | Bafilomycin | 8 | - | - | - | - | - | - | - | - | AJSZ01000001 |
| Streptomyces fungicidicus TXX3120 | Enduracidin | 7 | - | - | - | - | - | - | - | - | NZ_CP023407 |
| Streptomyces ghanaensis ATCC 14762 | Bafilomycin | 7 | - | - | - | - | - | - | - | - | DS999641 |
| Streptomyces pluripotens MUSC 135 | Antimicin | 7 | - | - | - | - | - | - | - | - | NZ_CP021080 |
| Frankia casuarinae CcI3 | Frankiamicin | 5 | - | - | - | - | - | - | - | - | NC_007777 |
Based on these findings, we devised a genome sequence-based in silico screening strategy for the identification of potential PSI producers, which was designated as protein synthesis inhibitor (‘psi’) target gene footprinting = Ψ-footprinting approach. The strategy involves the process of screening bacterial genome sequences with ARTS for the presence of a high number (≥20) of PSI hit genes and the abundance of RI genes (at least two), which allows for the identification of potential PSI producer strains. For dereplication purposes, the genome sequence of the identified strain is additionally analyzed with antiSMASH to sort out strains, which harbor BGCs encoding already known PSIs, allowing for the prioritization of strains likely to produce novel PSIs (Figure 4).
Figure 4.

Graphic illustration of the genome sequenced-based screening approach for the identification of PSI producer strains, designated Ψ-footprinting.
Verification of the Ψ-footprinting method by analyzing PSI producer strains of the Tübingen strain collection
To apply the Ψ-footprinting method to the previously discovered PSI producers of the Tübingen strain collection (5), the genomic DNA of the strains Tü 2108, Tü 2975, Tü 3180, Tü 6430, KNN 49.3e and A 4/2 was sequenced and analyzed as described above. A 4/2 was not further evaluated as it produced the eukaryotic PSI cycloheximide. Among the five strains, three (KNN 49.3e, Tü 3180 and Tü 6430) showed large numbers of hit genes, with abundances of 88, 50 and 45, respectively (Table 6). All strains harbored at least one RI gene, whereby for KNN 49.3e all seven RI genes were detected (Table 6). This showcase demonstrates that the Ψ-footprinting method enabled a genome sequence-based identification of three out of five PSI producer strains from the Tübingen strain collection.
Table 6.
List of protein synthesis inhibitor (PSI) producer strains from the Tübingen strain collection and their respective PSIs. Strains are ordered according to the occurrence of protein synthesis ‘hit genes’. The number of ‘resistance indicator’ (RI) genes is also listed
| Strain | Antibiotic (PSI) | Hit genes | RI genes |
|---|---|---|---|
| KNN 49.3e | Amicetin | 88 | 7 |
| Tü 3180 | Griseoviridin/Viridogrisein | 50 | 3 |
| Tü 6430 | Pactamycin | 45 | 3 |
| Tü 2975 | Pristinamycin | 16 | 1 |
| Tü 2108 | Berninamycin | 15 | 2 |
Ψ-footprinting method allows to prioritize potential PSI producers from the DSMZ strain collection
To apply the Ψ-footprinting approach to so far uncharacterized strains, the method was challenged with numerous genome-sequenced strains from the DSMZ strain collection. Here, the focus was on the analysis of genome sequences from rare actinomycetes in order to increase the probability of finding new substances. 406 actinomycetal genome sequences were examined. 118 genomes (30%) showed a high number of PSI hit genes (>20) (data not shown) and 141 (35%) strains had at least one RI gene (Supplementary Table S4). 110 (27%) of the strains met the two criteria for possible PSI production (≥20 PSI hit genes and at least two RI genes) and thus were candidate strains for PSI production. Genome sequences, which met these criteria, were analyzed with antiSMASH to exclude strains containing an already known PSI BGC. In total, 15 strains were prioritized for further analysis, which harbored up to 71 hit genes and five RI genes, as for example strain Jiangella alkaliphila DSM 45079 (Table 7).
Table 7.
List of possible protein synthesis inhibitor (PSI) producer strains of the DSMZ strain collection. Strains are ordered according to the occurrence of protein synthesis ‘hit genes’. Abundance of ‘resistance indicator’ (RI) genes/encoded proteins is highlighted as plus. Gene/protein absence is indicated as minus
| Species | DSM number | Hit gene | rpsE | rpsL | rplC | rplD | rplK | rplQ | rplV | RI genes |
|---|---|---|---|---|---|---|---|---|---|---|
| Jiangella alkaliphila | 45079 | 71 | + | + | - | + | - | + | + | 5 |
| Saccharopolyspora flava | 44771 | 64 | + | + | - | + | + | + | - | 5 |
| Actinokineospora cianjurensis | 45657 | 57 | + | + | - | + | + | - | - | 4 |
| Micromonospora aurantiaca | 43813 | 53 | - | - | - | + | + | + | + | 4 |
| Micromonospora violae | 45888 | 47 | - | - | - | + | - | + | + | 3 |
| Micromonospora purpureochromogenes | 43821 | 40 | + | + | - | - | - | - | + | 3 |
| Lentzea albidocapillata | 44073 | 40 | + | + | - | - | + | + | + | 5 |
| Nocardioides albertanoniae | 25218 | 39 | - | + | - | - | + | + | + | 4 |
| Actinomadura atramentaria | 43919 | 35 | - | + | - | + | - | + | + | 4 |
| Geodermatophilus nigrescens | 45408 | 34 | - | + | - | - | + | - | + | 3 |
| Hoyosella altamirensis | 45258 | 25 | + | + | - | + | - | - | + | 4 |
| Rhodococcus pyridinivorans | 44555 | 23 | + | - | - | + | - | - | + | 3 |
| Rhodococcus koreensis | 44498 | 22 | - | - | - | + | - | - | + | 2 |
| Williamsia marianensis | 44944 | 22 | + | - | - | + | - | - | + | 3 |
| Rhodococcus kroppenstedtii | 44908 | 20 | - | - | - | + | - | - | + | 2 |
To investigate whether the selected 15 strains produce PSIs, the strains were cultivated in three different cultivation media (NL800, MS and R5) shown to be suitable production media in previous studies (5). Culture samples were harvested at different time points (4, 7 and 10 days), extracted with ethyl acetate in a ratio of 1:1, concentrated 20-fold in vacuo and then redissolved in methanol. To investigate if the extracts contained substances with protein synthesis inhibiting activity, samples were analyzed using a coupled in vitro transcription/translation (ivTT) assay (31). This cell-free assay uses the E. coli translational machinery to express a green fluorescent protein (GFP) from a plasmid (pET28a-eGFP), reporting transcription/translation activity by fluorescence emission analysis. First, the assay was validated for the analysis of culture extracts and it was found to tolerate 30 μl of the concentrated culture extracts in a total assay volume of 100 μl without significant interference. Changes in GFP expression were monitored by measuring the fluorescence at 485/520 nm excitation/emission with a Tecan Infinite M200 Pro device and analyzing the data with the i-control™ Microplate Reader Software 1.11. To determine the maximum fluorescence value (GFP fluorescence = 100%), the assay was performed without the addition of antibiotics. As positive controls for effective protein synthesis inhibition, pure tetracycline and apramycin were used, respectively. As negative control, media extracts of the respective culture media (NL800, MS and R5) were tested to exclude that the observed inhibitory effects were caused by media components. Furthermore, extract samples from the strain S. coelicolor M1146 (M1146) were analyzed as a negative control. M1146 is a genetically engineered strain with deleted BGCs and thus lacking any antibiotic production (51). M1146 extracts were analyzed to exclude that the observed inhibitory effects were caused by components from the cell metabolism of actinomycetes. IvTT conducted with the media extract samples as negative controls yielded 70% - 85% maximal GFP fluorescence values and thus had only a minor inhibiting effect on the assay (Figure 5). The same was observed for the culture extracts of M1146, for which the fluorescence values varied between 70% and 90% (Figure 5). The addition of 5 μl tetracycline (15 mg/ml) (tet15) and 5 μl of apramycin (50 mg/ml) (apra50) resulted in very low GFP fluorescence values of 13% and 12%, respectively, indicating a strong inhibition of the ivTT assay (Figure 5). Based on these control measurements, an internal threshold was set, according to which fluorescence values between 0% and 40% were considered to represent specific inhibition of the ivTT assay and values of 0–20% indicate strong inhibition of the assay.
Figure 5.
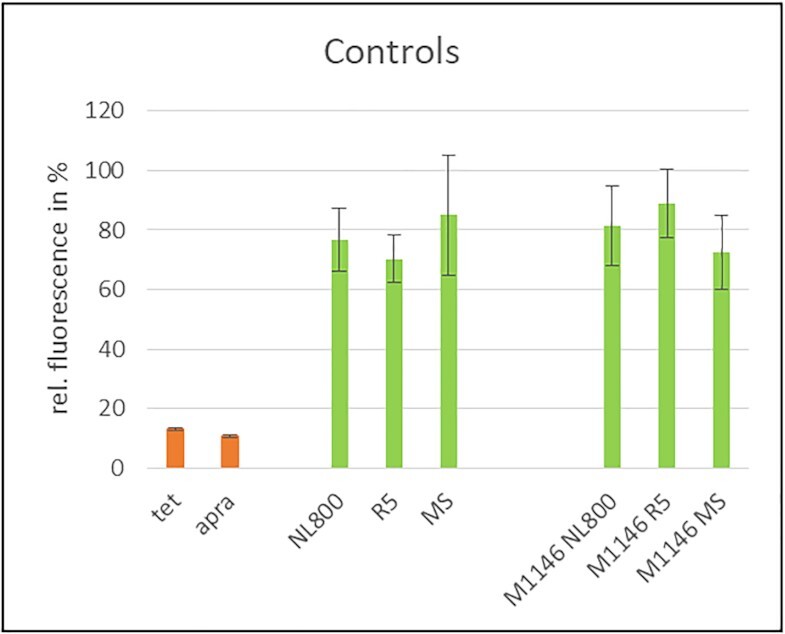
In vitro transcription/translation assay performed with culture extracts of controls. PSI antibiotics, tet15 and apra50 = positive control (orange), medium extracts, extracts of M1146 = negative control (green). For M1146, results are shown exemplary for samples from day 7. Measurements have been performed in triplicate using the same preparation of S12 extract.
IvTT assays with extract samples of strains DSM 44555, DSM 44498 and DSM 44908 showed no significant inhibition of GFP expression and thus the samples obviously did not contain protein synthesis inhibiting substances or at least they were not produced by the strains under the applied cultivation conditions (Supplementary Figure S4). IvTT assays with extract samples from the other twelve strains (DSM 44944, DSM 45888, DSM 45258, DSM 43821, DSM 44073, DSM 45079, DSM 45657, DSM 43813, DSM 43919, DSM 45408, DSM 25218 and DSM 44771) yielded significantly low fluorescence values ranging from 6% - 40% relative fluorescence (Supplementary Figure S5) and thus specific inhibition of the assay. This was observed for samples obtained from all tested media, whereby data are shown for the respective best production medium (Figure 6), while the complete dataset can be found in the supplemental material (Supplementary Figure S5). ivTT assays were additionally carried out with eleven culture extract sample from random strains of the Tübingen strain collection, of which none led to a specific inhibition (<40% fluorescence) of the assay, demonstrating that none of the strains produced a substance with PSI activity (Supplementary Figure S6).
Figure 6.

In vitro transcription/translation assay performed with culture extracts in NL800, MS and R5 media from 4, 7 and 10 days (optimal production time of each strain). PSI antibiotics, tet15 and apra50 = positive control (orange), medium extracts, extracts of M1146 = negative control (green), and extracts of DSM cultures (blue). Displayed are the extracts of each DSM strain that resulted in the most decrease of GFP production of the ivTT assay. Measurements have been performed in triplicate using the same preparation of S12 extract.
IvTT assays with culture extracts of DSM 43813, DSM 45079, DSM 45408, DSM 25218 and DSM 44771 grown in R5 resulted in the lowest fluorescence values (<20% maximal fluorescence), which indicated strong inhibition of the GFP expression, representatively shown for sample DSM 43813 (Figure 6, Supplementary Figure S5). Samples of strains DSM 43813 and DSM 45079 from sampling time points at days 7 and 10, respectively, reduced fluorescence to values of 17% and 12%, respectively (Figure 6). Samples of strains DSM 45408, DSM 25218 and DSM 44771 showed the strongest inhibiting effect for sampling time points at days 4, 7 and 10 with residual fluorescence values of 3%, 6% and 7%, respectively (Figure 6, Supplementary Figure S5). Since culture extracts of these five strains specifically inhibited the ivTT assay, it could be concluded that they interfere with protein expression and may contain a PSI. Thus, using the Ψ-footprinting-based prioritization of actinomycetes strains, potential PSI production was detected by ivTT assays for twelve out of the 15 (80%) selected DSM strains.
Identification of PSI substances from Ψ-footprinting-prioritized strains of the DSMZ strain collection
In order to identify the PSIs produced by the selected and analyzed DSM strains, the culture extracts were purified by semi-preparative HPLC followed by HPLC-MS analysis of the pre-purified substances. For these analyses, we focused on samples that showed the strongest inhibition of the ivTT assays, which included DSM 43813, DSM 45079, DSM 45408, DSM 25218 and DSM 44771. Methanolic culture extracts were prepared from samples harvested at the optimal production time point as reported above and protein synthesis inhibiting activity was verified in ivTT assays. For compound purification, semi-preparative HPLC was performed with the culture extracts. The fractions were collected in time-based mode every minute for 25 min and were analyzed for PSI activity via ivTT. Assay analysis revealed that individual purified fractions from all five DSM strain samples (DSM 43813, DSM 45079, DSM 45408, DSM 25218 and DSM 44771) showed significant inhibition of the ivTT assay (Figure 7, Supplementary Figures S7-S10). Interestingly, for all five extracts, fraction 8 (F8) showed the strongest inhibition, with fluorescence values of 9%, 30%, 4%, 33% and 7%, respectively (Figure 7, Supplementary Figures S7-S10), which suggested that F8 contained a protein synthesis inhibiting substance. HPLC-MS analysis of all extract samples and F8 fractions was performed to identify the respective PSI. The HPLC chromatograms of the extract samples of F8 of DSM 43813, DSM 45408, DSM 25218, and DSM 44771 of F8 showed a prominent peak at a retention time (RT) 7.2 min, as representatively shown for the sample DSM 25218 (Figure 8A). HPLC-MS analysis of the F8 samples of DSM 43813, DSM 45408, DSM 25218 and DSM 44771 revealed one distinct peak in positive and negative mode at RT 7.2 min (Figure 8B and C). This peak showed a characteristic UV–Vis spectrum with three λmax at 206 , 248 and 314 nm, respectively (Figure 8D). The peak was detected in positive mode at m/z = 424.2 [M + H]+ and in negative mode at m/z = 422.1 [M–H]– (Figure 8B and C). The mass of m/z = 423 is consistent with the mass of amicoumacin A (molecular weight 423.5; molecular formula C20H29N3O7) (52). The peak at RT 7.4 shows a prominent shoulder (Figure 8A). This often results due to the presence of two similar substances, which were not separated chromatographically. The shoulder showed a UV–Vis spectrum similar to that of amicoumacin A (Fig. 8G versus D). The UV–Vis spectrum of the peak was compared with an internal database, matching the entry of amicoumacin B (Figure 8G). This peak was detected in positive mode at m/z = 425.1 [M + H]+ and in negative mode at m/z = 423.1 [M–H]– (Figure 8E and F). The mass of m/z = 424 is consistent with the mass of amicoumacin B (molecular weight 424.4; molecular formula C20H28N2O8) (52). Amicoumacin A and B production have further been confirmed by high-resolution MS/MS (HRMS) analysis with F8 sample of DSM 45408, representatively (Supplementary Figure S11). The data obtained in this study (UV–Vis spectrum in combination with RT and mass of the peaks) showed that F8 of the samples DSM 43813, DSM 45408, DSM 25218 and DSM 44771 contained the PSIs amicoumacin A and B. Regarding F8 of strain DSM 45079, the mass of amicoumacin was not detected in this sample, and no mass or UV–Vis spectra of known PSIs could be assigned. It cannot be ruled out that amicoumacin was present in this sample in a very low concentration, which would fit to the lower inhibitory effect in the ivTT assay. On the other hand, this result could also indicate that the strain produces a novel PSI, but further cultivation and purification steps are necessary to allow accurate conclusions.
Figure 7.

In vitro transcription/translation assay performed with R5 culture extract of DSM 43813 from day 7. Displayed are the fractions generated by semi-preparative HPLC (blue). PSI antibiotic apra50 = positive control (orange). Fraction having the greatest inhibition is shown in red. Measurements have been performed in triplicate using the same preparation of S12 extract.
Figure 8.

(A) HPLC chromatogram from fraction F8 of the R5 extract from DSM 25218. Wavelength monitoring was performed at 260 nm. Arrows indicate amicoumacin A and B specific peaks at RT 7.2 min and 7.4 min, respectively. (B, C) Mass spectra from fraction F8 of the R5 extract of the amicoumacin producer strains. Amicoumacin A peaks in positive mode (B) m/z = 424.2 [M + H]+ and negative mode (C) m/z = 422.1 [M-H]– at RT 7.2 min are marked with red ellipses. (D) UV–Vis spectrum of DSM 25218 extract sample of the main peak at RT 7.2 min with amicoumacin B as reference and structure. Here, amicoumacin B was used as a reference since it was the only amicoumacin compound available in the internal database. (E, F) Mass spectra from fraction F8 of the R5 extract of the amicoumacin producer strains. Amicoumacin B peaks in positive mode (E) m/z = 425.1 [M + H]+ and negative mode (F) m/z = 423.1 [M-H]– at RT 7.4 min are marked with red ellipses. (G) UV–Vis spectrum of DSM 25218 extract sample of the shoulder peak at RT 7.4 min with amicoumacin B as reference and structure. The background was subtracted due to the overexposed masses of amicoumacin A for B/C and E/F (from main peak).
Amicoumacins are known PSIs, belonging to a class of isocoumarin antibiotics with antibacterial, antifungal, anticancer, and anti-inflammatory activities (53). They also show potent antibacterial activities against clinically relevant bacterial pathogens, such as Helicobacter pylori and methicillin-resistant Staphylococcus aureus (54,55). Amicoumacin affects bacterial growth by binding to the 30S subunit of the bacterial ribosome, specifically at the E-site codon of the mRNA, and thereby inhibits the translation process. This position is a rather unusual target site and implies a unique way of translation inhibition for amicoumacin (56). Amicoumacin has been reported as a natural product from different microbial producers, involving Gram-positive bacteria, such as certain representatives of Bacillus and Nocardia (54,57,58) but also the Gram-negative producer Xenorhabdus bovienii (53). In addition, our data revealed that amicoumacin is produced by various types of actinomycetes, namely Saccharopolyspora flava DSM 44771, Micromonospora aurantiaca DSM 43813, Nocardioides albertanoniae DSM 25218, and Geodermatophilus nigrescens DSM 45408, which belong to different orders of the phylum Actinobacteria, such as Pseudonocardiales, Micromonosporales, Propionobacteriales, and Geodermatophilales, respectively (Supplementary Figure S12). Thus, the ability to produce amicoumacin seems to be quite conserved across different groups of bacterial organisms, hinting at a so far unknown general biological function. The reason why we have not been able to dereplicate the substance based on BGC predictions is that the respective amicoumacin BGCs could not be identified by antiSMASH analysis. The amicoumacin BGC was described as nonribosomal peptide-polyketide hybrid BGC for the producer strain Xenorhabdus bovieni (53). However, the BGC is not part of the MIBiG repository and thus is not accessible as a reference in the antiSMASH database, which is why the BGCs were not identified by antiSMASH analysis for the amicoumacin producers of the DSMZ strain collection. Remarkably, also detailed manual sequence analyses did not lead to the identification of the amicoumacin BGCs. We therefore assume that the actinobacterial amicoumacin biosynthesis (high GC content bacteria) proceeds differently and is encoded by a strikingly different type of BGC than in the known producers of Xenorhabdus and Bacillus (low GC content bacteria). If so, this would speak for a convergent evolution of the amicoumacin BGCs in the phylogenetically distantly related producer strains. The fact that we have detected amicoumacin as PSI in our samples underscores the feasibility and reliability of the Ψ-footprinting strategy. The advantage of this method is that it is purely in silico-based. Conventional antibiotic screening approaches are generally much more labor-intensive since they involve screening a large number of strains, followed by extensive compound purifications and chemical analytics with the risk that the identified bioactive substance is already known. The Ψ-footprinting strategy is a fast and efficient method to specifically prioritize potential producers of PSIs based on genome-sequence information and thereby speeds up the lead discovery process. Until now, it was not possible to transfer this method to antibiotics with a different mode of action. The derived in silico strategy is so far only applicable for the prioritization of potential PSI producer strains, due to the number of core genes used as a reference set in the ARTS analysis. For the metabolic function of protein synthesis ∼100 core genes are defined in the reference set, which are significantly lower core gene numbers for other metabolic functions, such as DNA metabolism- or cell envelope-associated functions (16). Certainly, not all PSI producers can be detected with the Ψ-footprinting approach, as for example shown for Streptomyces griseus, a known producer of the PSI streptomycin (Table 4). Another limitation of the approach might be that it employs known RI genes as indicator genes, which could prevent the discovery of really novel PSIs with different MoAs. In this context, however, it would be possible to adopt a less stringent Ψ-footprinting run and exclude RI genes as a selection criterion, which would allow for a more explorative screening attempt. However, following the defined criteria for Ψ-footprinting, which are (i) a high number (≥20) of PSI hit genes and (ii) the additional abundance of RI genes, allows a reliable pre-selection of potential PSI producers. This was proven by the identification of twelve PSI producer strains from the 15 Ψ-selected strains in the current study, which inhibited protein synthesis in ivTT assays. In addition, the responsible PSI substances (amicoumacin A and B) were identified for four of these strains. Prioritized strains for which protein synthesis inhibiting activities were observed but no PSI identified yet are subject to ongoing work.
DATA AVAILABILITY
Genome sequencing data can be found at the National Center for Biotechnology Information (NCBI) linked to the accession numbers CAKMXI010000001-CAKMXI010000042 (Tü 2108), JAKGSH010000001-JAKGSH010000010 (Tü 6430), CALNVW010000001-CALNVW010000144 (A 4/2) and CP091196 (KNN 49.3e).
Supplementary Material
ACKNOWLEDGEMENTS
We thank Regina Ort-Winklbauer for excellent technical assistance and acknowledge additional experimental help from Oliver Hennrich and Meike Döppner with ivTT assays of control strains. We thank Imen Nouioui, Marlen Jando, Katharina Huber-Fischer and Carolin Pilke for supplying us with strains from the DSMZ culture collection. We are grateful to Nadine Ziemert for valuable support and discussions on the ARTS tool and thank Chambers Hughes for HRMS data on amicoumacin. We thank Mervyn Bibb from the John Innes Centre, UK for kindly providing strain S. coelicolor M1146.
Author contributions: Y.M. supervised and coordinated the study. K.W.W. carried out the agar-based bioreporter assays. F.H. cultivated the natural compound producers and performed purification of PSIs. A.K. and F.H. conducted HPLC–MS and HRMS analyses and dereplication of the antimicrobial compounds. J.S.S. performed the dereplication process of KNN 49.3e. R.O.W. isolated genomic DNA of PSI producers. A.W., D.W. and J.K. provided the sequence data for strain Tü 2108 and A 4/2. F.H. and Y.M. carried out genome mining approaches. F.H. performed the phylogenomic analysis. F.H. and Y.M. developed the Ψ-footprinting method. FH established and applicated the Ψ-footprinting method. F.H. established the assay conditions for the ivTT assay and performed all experiments. F.H. and Y.M. wrote the manuscript. All authors discussed results, contributed to the article and approved the submitted version.
Contributor Information
Franziska Handel, Department of Microbiology/Biotechnology, Interfaculty Institute of Microbiology and Infection Medicine, Faculty of Science, University of Tübingen, Auf der Morgenstelle 28, 72076 Tübingen, Germany; German Center for Infection Research (DZIF), Partner Site Tübingen, Tübingen, Germany.
Andreas Kulik, Department of Microbiology/Biotechnology, Interfaculty Institute of Microbiology and Infection Medicine, Faculty of Science, University of Tübingen, Auf der Morgenstelle 28, 72076 Tübingen, Germany; Department of Microbial Bioactive Compounds; Interfaculty Institute of Microbiology and Infection Medicine, University of Tübingen; Tübingen, Baden-Württemberg 72076, Germany.
Katharina W Wex, Department of Microbial Bioactive Compounds; Interfaculty Institute of Microbiology and Infection Medicine, University of Tübingen; Tübingen, Baden-Württemberg 72076, Germany; German Center for Infection Research (DZIF), Partner Site Tübingen, Tübingen, Germany.
Anne Berscheid, Department of Microbial Bioactive Compounds; Interfaculty Institute of Microbiology and Infection Medicine, University of Tübingen; Tübingen, Baden-Württemberg 72076, Germany; German Center for Infection Research (DZIF), Partner Site Tübingen, Tübingen, Germany.
Julian S Saur, Biomolecular Chemistry, Institute of Organic Chemistry, University of Tübingen, Tübingen, Baden-Württemberg 72076, Germany.
Anika Winkler, Center for Biotechnology (CeBiTec), Bielefeld University, Universitätsstraße 27, 33615 Bielefeld, Germany.
Daniel Wibberg, Center for Biotechnology (CeBiTec), Bielefeld University, Universitätsstraße 27, 33615 Bielefeld, Germany.
Jörn Kalinowski, Center for Biotechnology (CeBiTec), Bielefeld University, Universitätsstraße 27, 33615 Bielefeld, Germany.
Heike Brötz-Oesterhelt, Department of Microbial Bioactive Compounds; Interfaculty Institute of Microbiology and Infection Medicine, University of Tübingen; Tübingen, Baden-Württemberg 72076, Germany; German Center for Infection Research (DZIF), Partner Site Tübingen, Tübingen, Germany; Cluster of Excellence Controlling Microbes to Fight Infection, Germany.
Yvonne Mast, Department of Microbiology/Biotechnology, Interfaculty Institute of Microbiology and Infection Medicine, Faculty of Science, University of Tübingen, Auf der Morgenstelle 28, 72076 Tübingen, Germany; German Center for Infection Research (DZIF), Partner Site Tübingen, Tübingen, Germany; Department Bioresources for Bioeconomy and Health Research, Leibniz Institute DSMZ - German Collection of Microorganisms and Cell Cultures, Inhoffenstraße 7B, 38124 Braunschweig, Germany; Technical University Braunschweig, Department of Microbiology, Rebenring 56, 38106 Braunschweig, Germany.
SUPPLEMENTARY DATA
Supplementary Data are available at NARGAB Online.
FUNDING
German Center for Infection Research (DZIF) [TTU 09.819, TTU 09.812]; H.B.-Ö. is also grateful for infrastructural funding by the Cluster of Excellence Controlling Microbes to Fight Infection [EXC 2124, 390838134].
Conflict of interest statement. The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
REFERENCES
- 1. WHO Antibiotic Resistance: Multi-country Public Awareness Survey. 2015; World Health Organization. [Google Scholar]
- 2. Lewis K. Platforms for antibiotic discovery. Nat. Rev. Drug Discov. 2013; 12:371–387. [DOI] [PubMed] [Google Scholar]
- 3. Wright G.D. Opportunities for natural products in 21st century antibiotic discovery. Nat. Prod. Rep. 2017; 34:694–701. [DOI] [PubMed] [Google Scholar]
- 4. Wencewicz T.A. New antibiotics from Nature's chemical inventory. Bioorganic Med. Chem. 2016; 24:6227–6252. [DOI] [PubMed] [Google Scholar]
- 5. Wex K.W., Saur J.S., Handel F., Ortlieb N., Mokeev V., Kulik A., Niedermeyer T.H.J., Mast Y., Grond S., Berscheid A.et al.. Bioreporters for direct mode of action-informed screening of antibiotic producer strains. Cell Chem. Biol. 2021; 28:1242–1252. [DOI] [PubMed] [Google Scholar]
- 6. Ortlieb N., Klenk E., Kulik A., Niedermeyer T.H.J.. Development of an agar-plug cultivation system for bioactivity assays of actinomycete strain collections. PLoS One. 2021; 16:e0258934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Scherlach K., Hertweck C.. Mining and unearthing hidden biosynthetic potential. Nat. Commun. 2021; 12:3864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Tanaka Y., Omura S.. Metabolism and products of actinomycetes. An introduction. Actinomycetologica. 1990; 4:13–14. [Google Scholar]
- 9. Bérdy J. Bioactive microbial metabolites. J. Antibiot. (Tokyo). 2005; 58:1–26. [DOI] [PubMed] [Google Scholar]
- 10. Carrano L., Marinelli F.. The relevance of chemical dereplication in microbial natural product screening. J. Appl. Bioanal. 2015; 1:55–67. [Google Scholar]
- 11. Gavriilidou A., Kautsar S.A., Zaburannyi N., Krug D., Müller R., Medema M.H., Ziemert N.. ACompendium of specialized metabolite biosynthetic diversity encoded in bacterial genomes. Nat. Microbiol. 2022; 7:726–735. [DOI] [PubMed] [Google Scholar]
- 12. Medema M.H., Kottmann R., Yilmaz P., Cummings M., Biggins J.B., Blin K., de Bruijn I., Chooi Y.H., Claesen J., Coates R.C.et al.. Minimum information about a biosynthetic gene cluster. Nat. Chem. Biol. 2015; 11:625–631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Belknap K.C., Park C.J., Barth B.M., Andam C.P.. Genome mining of biosynthetic and chemotherapeutic gene clusters in Streptomyces bacteria. Sci. Rep. 2020; 10:2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Baltz R.H. Gifted microbes for genome mining and natural product discovery. J. Ind. Microbiol. Biotechnol. 2017; 44:573–588. [DOI] [PubMed] [Google Scholar]
- 15. Blin K., Shaw S., Kloosterman A.M., Charlop-Powers Z., van Wezel G.P., Medema M.H., Weber T.. antiSMASH 6.0: improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021; 49:W29–W35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Alanjary M., Kronmiller B., Adamek M., Blin K., Weber T., Huson D., Philmus B., Ziemert N.. The Antibiotic Resistant Target Seeker (ARTS), an exploration engine for antibiotic cluster prioritization and novel drug target discovery. Nucleic Acids Res. 2017; 45:W42–W48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Baumann S., Herrmann J., Raju R., Steinmetz H., Mohr K.I., Hüttel S., Harmrolfs K., Stadler M., Müller R.. Cystobactamids: myxobacterial topoisomerase inhibitors exhibiting potent antibacterial. Angew. Chem. Int. Ed. 2014; 53:14605–14609. [DOI] [PubMed] [Google Scholar]
- 18. Tran J.H., Jacoby G.A.. Mechanism of plasmid-mediated quinolone resistance. Proc. Natl. Acad. Sci. U.S.A. 2002; 99:5638–5642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Kale A.J., McGlinchey R.P., Lechner A., Moore B.S.. Bacterial self-resistance to the natural proteasome inhibitor salinosporamide A. ACS Chem. Biol. 2011; 6:1257–1264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Tang X., Li J., Millán-Aguiñaga N., Zhang J.J., O’Neill E.C., Ugalde J.A., Jensen P.R., Mantovani S.M., Moore B.S. Identification of thiotetronic acid antibiotic biosynthetic pathways by Target-directed genome mining. ACS Chem. Biol. 2015; 10:2841–2849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Mungan M.D., Alanjary M., Blin K., Weber T., Medema M.H., Ziemert N.. ARTS 2.0: feature updates and expansion of the antibiotic resistant target seeker for comparative genome mining. Nucleic Acids Res. 2020; 48:W546–W552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Poehlsgaard J., Douthwaite S.. The bacterial ribosome as a target for antibiotics. Nat. Rev. Microbiol. 2005; 3:870–881. [DOI] [PubMed] [Google Scholar]
- 23. Wilson D.N. Ribosome-targeting antibiotics and mechanisms of bacterial resistance. Nat. Rev. Microbiol. 2014; 12:35–48. [DOI] [PubMed] [Google Scholar]
- 24. Xu Z., Culver G.M.. Differential assembly of 16S rRNA domains during 30S subunit formation. RNA. 2010; 16:1990–2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Carter A.P., Clemons W.M., Brodersen D.E., Morgan-Warren R.J., Wimberly B.T., Ramakrishnan V.. Functional insights from the structure of the 30S ribosomal subunit and its interactions with antibiotics. Nature. 2000; 407:340–348. [DOI] [PubMed] [Google Scholar]
- 26. Peske F., Savelsbergh A., Katunin V.I., Rodnina M.V., Wintermeyer W.. Conformational changes of the small ribosomal subunit during elongation factor G-dependent tRNA-mRNA translocation. J. Mol. Biol. 2004; 343:1183–1194. [DOI] [PubMed] [Google Scholar]
- 27. Pape T., Wintermeyer W., Rodnina M.V.. Conformational switch in the decoding region of 16S rRNA during aminoacyl-tRNA selection on the ribosome. Nat. Struct. Biol. 2000; 7:104–107. [DOI] [PubMed] [Google Scholar]
- 28. Long K.S., Vester B.. Resistance to linezolid caused by modifications at its binding site on the ribosome. Antimicrob. Agents Chemother. 2012; 56:603–612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Polikanov Y.S., Szal T., Jiang F., Gupta P., Matsuda R., Shiozuka M., Steitz T.A., Vázquez-Laslop N., Mankin A.S.. Negamycin interferes with decoding and translocation by simultaneous interaction with rRNA and tRNA. Mol. Cell. 2014; 56:541–550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Belova L., Tenson T., Xiong L., McNicholas P.M., Mankin A.S.. A novel site of antibiotic action in the ribosome: interaction of evernimicin with the large ribosomal subunit. Proc. Natl. Acad. Sci. U.S.A. 2001; 98:3726–3731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Musiol-Kroll E.M., Zubeil F., Schafhauser T., Härtner T., Kulik A., McArthur J., Koryakina I., Wohlleben W., Grond S., Williams G.J.et al.. Polyketide bioderivatization using the promiscuous acyltransferase KirCII. ACS Synth. Biol. 2017; 6:421–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Kieser T., Bibb M.J., Buttner M.J., Chater K.F., Hopwood D.A.John Innes Foundation . Practical Streptomyces Genetics. Engl. 3. 2000; Norwich: John Innes Found. [Google Scholar]
- 33. Hennrich O., Handel F., Ort-Winklbauer R., Mast Y.. Genome sequences of two putative streptogramin producers, Streptomyces sp. Strains TÜ 2975 and TÜ 3180, from the tübingen strain collection. Microbiol. Resour. Announc. 2020; 9:e01582-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Zecher K., Suleiman M., Wibberg D., Winkler A., Philipp B., Kalinowski J.. Draft genome sequence of Donghicola sp. strain Karma, a model organism for monomethylamine-degrading nonmethylotrophic bacteria. Genome Announc. 2017; 5:e01623-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Meier-Kolthoff J.P., Göker M.. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019; 10:2182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Meier-Kolthoff J.P., Auch A.F., Klenk H.P., Göker M.. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics. 2013; 14:60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Medema M.H., Blin K., Cimermancic P., De Jager V., Zakrzewski P., Fischbach M.A., Weber T., Takano E., Breitling R.. AntiSMASH: rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011; 39:W339–W346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Krause J., Handayani I., Blin K., Kulik A., Mast Y.. Disclosing the potential of the SARP-Type regulator PapR2 for the activation of antibiotic gene clusters in streptomycetes. Front. Microbiol. 2020; 11:225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Fiedler H.-P. Biosynthetic capacities of actinomycetes. 1 screening for secondary metabolites by HPLC and UV-Visible absorbance spectral libraries. Nat. Prod. Lett. 1993; 2:119–128. [Google Scholar]
- 40. Taylor & Francis Group Dictionary of Natural Products 30. 2020; 1:CRC Press. [Google Scholar]
- 41. Ninomiya A., Kodani S.. Isolation and identification of berninamycin a from Streptomyces atroolivaceus. Asian J. Chem. 2013; 25:490–492. [Google Scholar]
- 42. Malcolmson S.J., Young T.S., Ruby J.G., Skewes-Cox P., Walsh C.T.. The posttranslational modification cascade to the thiopeptide berninamycin generates linear forms and altered macrocyclic scaffolds. Proc. Natl. Acad. Sci. U.S.A. 2013; 110:8483–8488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Thompson J., Cundliffe E., Stark M.J.R.. The mode of action of berninamycin and the mechanism of resistance in the producing organism, Streptomycesbernensis. J. Gen. Microbiol. 1982; 128:875–884. [DOI] [PubMed] [Google Scholar]
- 44. Bilgin N., Richter A.A., Ehrenberg M., Dahlberg A.E., Kurland C.G.. Ribosomal RNA and protein mutants resistant to spectinomycin. EMBO J. 1990; 9:735–739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Wilcox S.K., Cavey G.S., Pearson J.D.. Single ribosomal protein mutations in antibiotic-resistant bacteria analyzed by mass spectrometry. Antimicrob. Agents Chemother. 2001; 45:3046–3055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Klitgaard R.N., Ntokou E., Nørgaard K., Biltoft D., Hansen L.H., Trædholm N.M., Kongsted J., Vester B.. Mutations in the bacterial ribosomal protein L3 and their association with antibiotic resistance. Antimicrob. Agents Chemother. 2015; 59:3518–3528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Chittum H.S., Champney W.S.. Ribosomal protein gene sequence changes in erythromycin-resistant mutants of Escherichiacoli. J. Bacteriol. 1994; 176:6192–6198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Weisblum B. Erythromycin resistance by ribosome modification. Antimicrob. Agents Chemother. 1995; 39:577–585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Cameron D.M., Thompson J., Gregory S.T., March P.E., Dahlberg A.E.. Thiostrepton-resistant mutants of Thermusthermophilus. Nucleic Acids Res. 2004; 32:3220–3227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Tipper D.J., Johnson C.W., Ginther C.L., Leighton T., Wittmann H.G.. Erythromycin resistant mutations in Bacillussubtilis cause temperature sensitive sporulation. MGGMol. Gen. Genet. 1977; 150:147–159. [DOI] [PubMed] [Google Scholar]
- 51. Gomez-Escribano J.P., Bibb M.J.. Engineering Streptomycescoelicolor for heterologous expression of secondary metabolite gene clusters. Microb. Biotechnol. 2011; 4:207–215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Hashimoto M., Taguchi T., Nishida S., Ueno K., Koizumi K., Aburada M., Ichinose K., Hashimoto M., Taguchi T., Nishida S.et al.. Isolation of 8′-Phosphate ester derivatives of amicoumacins: structure-activity relationship of hydroxy amino acid moiety. J. Antibiot. (Tokyo). 2007; 60:752–756. [DOI] [PubMed] [Google Scholar]
- 53. Park H.B., Perez C.E., Perry E.K., Crawford J.M.. Activating and attenuating the amicoumacin antibiotics. Molecules. 2016; 21:824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Pinchuk I.V., Bressollier P., Verneuil B., Fenet B., Sorokulova I.B., Mégraud F., Urdaci M.C.. In vitro anti-Helicobacter pylori activity of the probiotic strain Bacillussubtilis 3 is due to secretion of antibiotics. Antimicrob. Agents Chemother. 2001; 45:3156–3161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Lama A., Pané-Farré J., Chon T., Wiersma A.M., Sit C.S., Vederas J.C., Hecker M., Nakano M.M.. Response of methicillin-resistant Staphylococcusaureus to Amicoumacin A. PLoS One. 2012; 7:e34037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Maksimova E.M., Vinogradova D.S., Osterman I.A., Kasatsky P.S., Nikonov O.S., Milón P., Dontsova O.A., Sergiev P.V., Paleskava A., Konevega A.L.. Multifaceted mechanism of amicoumacin a inhibition of bacterial translation. Front. Microbiol. 2021; 12:172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Itoh J., Omoto S., Shomura T., Nishizawa N., Miyado S., Yuda Y., Shibata U., Inouye S.. Amicoumacin a, a new antibiotic with strong antiinflammatory and antiulcer activity. J. Antibiot. (Tokyo). 1981; 34:611–613. [DOI] [PubMed] [Google Scholar]
- 58. Sun W., Zhang Y.Q., Huang Y., Zhang Y.Q., Yang Z.Y., Liu Z.H.. Nocardia jinanensis sp. nov., an amicoumacin B-producing actinomycete. Int. J. Syst. Evol. Microbiol. 2009; 59:417–420. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Genome sequencing data can be found at the National Center for Biotechnology Information (NCBI) linked to the accession numbers CAKMXI010000001-CAKMXI010000042 (Tü 2108), JAKGSH010000001-JAKGSH010000010 (Tü 6430), CALNVW010000001-CALNVW010000144 (A 4/2) and CP091196 (KNN 49.3e).