

Abstract
The comparative genomics of butterflies yields additional insights into their phylogeny and classification that are compiled here. As a result, 3 genera, 5 subgenera, 5 species, and 3 subspecies are proposed as new, i.e., in Hesperiidae: Antina Grishin, gen. n. (type species Antigonus minor O. Mielke, 1980), Pompe Grishin and Lamas, gen. n. (type species Lerema postpuncta Draudt, 1923), and Curva Grishin, gen. n. (type species Moeris hyagnis Godman, 1900); in Lycaenidae: Fussia Grishin, subgen. n. (type species Polyommatus standfussi Grum-Grshimailo, 1891) and Pava Grishin, subgen. n. (type species Thecla panava Westwood, 1852); in Hesperiidae: Monoca Grishin, subgen. n. (type species Tagiades monophthalma Plötz, 1884), Putuma Grishin, subgen. n. (type species Tisias putumayo Constantino and Salazar, 2013), and Rayia Grishin, subgen. n. (type species Mastor perigenes Godman, 1900); Cissia wahala Grishin, sp. n. (Nymphalidae; type locality in Mexico: Oaxaca); in Hesperiidae: Hedone mira Grishin and Lamas, sp. n. (type locality in Peru: Apurímac), Vidius pompeoides Grishin, sp. n. (type locality in Brazil: Amazonas), Parphorus hermieri Grishin, sp. n. (Hesperiidae; type locality in Brazil: Rondônia), and Zenis par Grishin, sp. n. (Hesperiidae; type locality in Peru: Cuzco); in Pieridae: Glutophrissa drusilla noroesta Grishin, ssp. n. (type locality in USA: Texas, Cameron Co.) and Pieris marginalis siblanca Grishin, ssp. n. (type locality in USA: New Mexico, Lincoln Co.), and Argynnis cybele neomexicana Grishin, ssp. n. (Nymphalidae; type locality in USA: New Mexico, Sandoval Co.). Acidalia leto valesinoides-alba Reuss, [1926] and Acidalia nokomis valesinoides-alba Reuss, [1926] are unavailable names. Neotypes are designated for Mylothris margarita Hübner, [1825] (type locality in Brazil) and Papilio coras Cramer, 1775 (type locality becomes USA: Pennsylvania, Montgomery Co., Flourtown). Mylothris margarita Hübner, [1825] becomes a junior objective synonym of Pieris ilaire Godart, 1819, currently a junior subjective synonym of Glutophrissa drusilla (Cramer, 1777). Lectotypes are designated for Hesperia ceramica Plötz, 1886 (type locality in Indonesia: Seram Island), Pamphila trebius Mabille, 1891 (type locality Colombia: Bogota), Methionopsis modestus Godman, 1901 and Papias microsema Godman, 1900 (type locality in Mexico: Tabasco), Hesperia fusca Grote & Robinson, 1867 (type locality in USA: Georgia), Goniloba corusca Herrich-Schäffer, 1869, and Goniloba devanes Herrich-Schäffer, 1869; the type localities of the last two species, together with Pamphila stigma Skinner, 1896 and Carystus (Argon) lota (Hewitson, 1877), are deduced to be in South America. Type locality of Junonia pacoma Grishin, 2020 is in Sinaloa, not Sonora (Mexico). Abdomen is excluded from the holotype of Staphylus ascalon (Staudinger, 1876). Furthermore, a number of taxonomic changes are proposed. Alciphronia Koçak, 1992 is treated as a subgenus, not a synonym of Heodes Dalman, 1816. The following genera are treated as subgenera: Lafron Grishin, 2020 of Lycaena [Fabricius], 1807, Aremfoxia Real, 1971 of Epityches D’Almeida, 1938, Placidina D’Almeida, 1928 of Pagyris Boisduval, 1870, and Methionopsis Godman, 1901 of Mnasinous Godman, 1900. Polites (Polites) coras (Cramer, 1775) is not a nomen dubium but a valid species. The following are species-level taxa (not subspecies or synonyms of taxa given in parenthesis): Lycaena pseudophlaeas (Lucas, 1866) and Lycaena hypophlaeas (Boisduval, 1852) (not Lycaena phlaeas (Linnaeus, 1761), Satyrium dryope (W. H. Edwards, 1870) (not Satyrium sylvinus (Boisduval, 1852)), Apodemia cleis (W. H. Edwards, 1882) (not Apodemia zela (Butler, 1870)), Epityches thyridiana (Haensch, 1909), comb. nov. (not Epityches ferra Haensch, 1909, comb. nov.), Argynnis bischoffii W. H. Edwards, 1870 (not Argynnis mormonia Boisduval, 1869), Argynnis leto Behr, 1862 (not Argynnis cybele (Fabricius, 1775)), Boloria myrina (Cramer, 1777) (not Boloria selene ([Denis & Schiffermüller], 1775)), Phyciodes jalapeno J. Scott, 1998 (not Phyciodes phaon (W. H. Edwards, 1864)), Phyciodes incognitus Gatrelle, 2004 and Phyciodes diminutor J. Scott, 1998 (not Phyciodes cocyta (Cramer, 1777)), Phyciodes orantain J. Scott, 1998 (not Phyciodes tharos (Drury, 1773)), Phyciodes anasazi J. Scott, 1994 (not Phyciodes batesii (Reakirt, [1866])), Cercyonis silvestris (W. H. Edwards, 1861) (not Cercyonis sthenele (Boisduval, 1852)), Paramacera allyni L. Miller, 1972 and Paramacera rubrosuffusa L. Miller, 1972 (not Paramacera xicaque (Reakirt, [1867])), Cissia cheneyorum (R. Chermock, 1949), Cissia pseudocleophes (L. Miller, 1976), and Cissia anabelae (L. Miller, 1976) (not Cissia rubricata (W. H. Edwards, 1871)), Tarsoctenus gaudialis (Hewitson, 1876) (not Tarsoctenus corytus (Cramer, 1777)), Nisoniades inca (Lindsey, 1925) (not Nisoniades mimas (Cramer, 1775), Xenophanes ruatanensis Godman & Salvin, 1895 (not Xenophanes tryxus (Stoll, 1780)), Lotongus shigeoi Treadaway & Nuyda, 1994, Lotongus balta Evans, 1949, Lotongus zalates (Mabille, 1893), and Lotongus taprobanus (Plötz, 1885) (not Lotongus calathus (Hewitson, 1876)), Oxynthes martius (Mabille, 1889) (not Oxynthes corusca (Herrich-Schäffer, 1869)), Notamblyscirtes durango J. Scott, 2017 (not Notamblyscirtes simius W. H. Edwards, 1881), Hedone praeceps Scudder, 1872, Hedone catilina (Plötz, 1886), and Hedone calla (Evans, 1955) (not Hedone vibex (Geyer, 1832)), Atalopedes huron (W. H. Edwards, 1863) (not Atalopedes campestris (Boisduval, 1852)), Papias microsema Godman, 1900 (not Mnasinous phaeomelas (Hübner, [1829]), comb. nov.), Papias unicolor (Hayward, 1938) and Papias monus Bell, 1942 (not Papias phainis Godman, 1900), Nastra leuconoides (Lindsey, 1925) (not Nastra leucone (Godman, 1900)), Nastra fusca (Grote & Robinson, 1867) (not Nastra lherminier (Latreille, [1824])), Zenis hemizona (Dyar, 1918) and Zenis janka Evans, 1955 (not Zenis jebus (Plötz, 1882)), Carystus (Argon) argus Möschler, 1879 (not Carystus (Argon) lota Hewitson, 1877), and Lycas devanes (Herrich-Schäffer, 1869) (not Lycas argentea (Hewitson, 1866)). Borbo impar ceramica (Plötz, 1886), comb. nov. is not a synonym of Pelopidas agna larika (Pagenstecher, 1884) but a valid subspecies. Parnassius smintheus behrii W. H. Edwards, 1870 and Cercyonis silvestris incognita J. Emmel, T. Emmel & Mattoon, 2012 are subspecies, not species. The following are junior subjective synonyms: Shijimiaeoides Beuret, 1958 of Glaucopsyche Scudder, 1872, Micropsyche Mattoni, 1981 of Turanana Bethune-Baker, 1916, Cyclyrius Butler, 1897 of Leptotes Scudder, 1876, Mesenopsis Godman & Salvin, 1886 of Xynias Hewitson, 1874, Carystus tetragraphus Mabille, 1891 of Lotongus calathus parthenope (Plötz, 1886), Parnara bipunctata Elwes & J. Edwards, 1897 of Borbo impar ceramica (Plötz, 1886), Hesperia peckius W. Kirby, 1837 of Polites (Polites) coras (Cramer, 1775), and Lerodea neamathla Skinner & R. Williams, 1923 of Nastra fusca (Grote & Robinson, 1867). The following transfers are proposed: of species between genera (i.e., revised genus-species combinations): Nervia niveostriga (Trimen, 1864) (not Kedestes Watson, 1893), Leona lota Evans, 1937 (not Lennia Grishin, 2022), Leona pruna (Evans, 1937) and Leona reali (Berger, 1962) (not Pteroteinon Watson, 1893), Mnasinous phaeomelas (Hübner, [1829]) (not Papias Godman, 1900), Saturnus jaguar (Steinhauser, 2008) (not Parphorus Godman, 1900), Parphorus harpe (Steinhauser, 2008) (not Saturnus Evans, 1955), Parphorus kadeni (Evans, 1955) (not Lento Evans, 1955), and Calpodes chocoensis (Salazar & Constantino, 2013) (not Megaleas Godman, 1901); of subspecies between species (i.e., revised species-subspecies combinations): Melitaea sterope W. H. Edwards, 1870 of Chlosyne palla (Boisduval, 1852) (not Chlosyne acastus (W. H. Edwards, 1874)) and Panoquina ocola distipuncta Johnson & Matusik, 1988 of Panoquina lucas (Fabricius, 1793); and junior subjective synonym transferred between species: Rhinthon zaba Strand, 1921 of Conga chydaea (A. Butler, 1877), not Cynea cynea (Hewitson, 1876), Pamphila stigma Skinner, 1896 of Hedone catilina (Plötz, 1886), not Hedone praeceps Scudder, 1872, and Pamphila ortygia Möschler, 1883 of Panoquina hecebolus (Scudder, 1872), not Panoquina ocola (W. H. Edwards, 1863). Proposed taxonomic changes result in additional revised species-subspecies combinations: Lycaena pseudophlaeas abbottii (Holland, 1892), Satyrium dryope putnami (Hy. Edwards, 1877), Satyrium dryope megapallidum Austin, 1998, Satyrium dryope itys (W. H. Edwards, 1882), Satyrium dryope desertorum (F. Grinnell, 1917), Argynnis bischoffi opis W. H. Edwards, 1874, Argynnis bischoffi washingtonia W. Barnes & McDunnough, 1913, Argynnis bischoffi erinna W. H. Edwards, 1883, Argynnis bischoffi kimimela Marrone, Spomer & J. Scott, 2008, Argynnis bischoffi eurynome W. H. Edwards, 1872, Argynnis bischoffi artonis W. H. Edwards, 1881, Argynnis bischoffi luski W. Barnes & McDunnough, 1913, Argynnis leto letona (dos Passos & Grey, 1945), Argynnis leto pugetensis (F. Chermock & Frechin, 1947), Argynnis leto eileenae (J. Emmel, T. Emmel & Mattoon, 1998), Boloria myrina nebraskensis (W. Holland, 1928), Boloria myrina sabulocollis Kohler, 1977, Boloria myrina tollandensis (W. Barnes & Benjamin, 1925), Boloria myrina albequina (W. Holland, 1928), Boloria myrina atrocostalis (Huard, 1927), Boloria myrina terraenovae (W. Holland, 1928), Phyciodes anasazi apsaalooke J. Scott, 1994, Polites coras surllano J. Scott, 2006, and Curva darienensis (Gaviria, Siewert, Mielke & Casagrande, 2018). Specimen curated as the holotype of Acidalia leto valesinoides-alba Reuss, [1926] is Argynnis leto letona (dos Passos & Grey, 1945) (not A. leto leto Behr, 1862) from USA: Utah, Provo. A synonymic list of available genus-group names for Lycaeninae [Leach], [1815] is given. Unless stated otherwise, all subgenera, species, subspecies and synonyms of mentioned genera and species are transferred with their parent taxa, and others remain as previously classified.
Keywords: nomenclature, taxonomy, classification, genomics, phylogeny, biodiversity
INTRODUCTION
In this study, we continue the exploration of the phylogenetic classification of butterflies aided by genomic sequencing. The general philosophy, strategy, and details of the methods follow our previous publications (Cong et al. 2019a, b; Li et al. 2019; Zhang et al. 2019a, b, c, d; Cong et al. 2020; Zhang et al. 2020; Cong et al. 2021; Zhang et al. 2021; Robbins et al. 2022; Zhang et al. 2022b, c). Here, we report further findings that are encountered as whole genomic shotgun datasets for additional specimens are being assembled and comparatively analyzed. We place emphasis on the sequencing of primary type specimens that provide objective references for the names (Zhang et al. 2022a). When type localities are unknown, we deduce them by genomic comparison of the type specimens with specimens from known localities (Cong et al. 2021). Criteria used for genera, subgenera, species, and subspecies are the same as we employed and discussed previously (Cong et al. 2019a, b; Li et al. 2019; Zhang et al. 2019b, d; Cong et al. 2020; Zhang et al. 2020; Zhang et al. 2021; Zhang et al. 2022b).
Because speciation and extinction patterns are linked to geological events simultaneously affecting many phylogenetic lineages, we observe levels in phylogenetic trees, i.e., periods of rapid diversification followed by the reduced number of splits that result in longer internal branches at about the same distance from the root (or the leaves). Genera are defined as the most prominent level in genomic trees between tribes and species that mostly corresponds to the current classification into genera. Subgenera form a rather prominent level between genera and species. Species are delineated by a combination of criteria that include genetic differentiation in the Z chromosome measured by Fst (>0.25 typically corresponds to distinct species) and gene exchange Gmin (<0.7 for distinct species) (Cong et al. 2019a), COI barcode difference (usually >2% for distinct species) (Hebert et al. 2003) and its correlation with phenotypic differences (Lukhtanov et al. 2016), and the prominence of species-level clades (Zhang et al. 2022c).
Diagnostic DNA characters are given as abbreviations for either of the three reference genomes: Pieris rapae (Linnaeus, 1758) (pra) (Shen et al. 2016), Calycopis cecrops (Fabricius, 1793) (cce) (Cong et al. 2016), or Cecropterus lyciades (Geyer, 1832) (aly) (Shen et al. 2017), and for the COI barcode: e.g., aly728.44.1:G672C means position 672 in exon 1 of gene 44 from scaffold 728 of C. lyciades (formerly in Achalarus Scudder, 1872, thus aly; cce would be for C. cecrops; no prefix and : for COI barcode) reference genome is C, changed from G in the ancestor.
The sections below follow the standardized format. Taxonomic act is given as the title. For cited genera and subgenera, type species are listed in parenthesis. Type localities are specified. Sections are illustrated by a segment of a nuclear genomic tree (or the Z chromosome tree when specified) with species necessary to support the conclusion. Currently employed names and combinations (Lamas 2004; Mielke 2005; Pelham 2008) are used in the figures, including recently proposed changes (Cong et al. 2019b; Zhang et al. 2019b; Zhang et al. 2020; Zhang et al. 2021; Pelham 2022; Zhang et al. 2022b). New combinations and taxonomic changes are given in the text and figure legends. The sections are ordered by family and generally in their taxonomic order deduced from genome-scale phylogeny complemented by phenotypic considerations. Whole genome shotgun datasets we obtained and used in this work are available from the NCBI database <https://www.ncbi.nlm.nih.gov/> as BioProject PRJNA883758, and BioSample entries of the project contain the locality and other collection data of the sequenced specimens shown in the trees. COI barcode sequences have been deposited in GenBank with accessions OP231464-OP231472, OP323110-OP323113, and OP381659-OP381661. Exon sequences with diagnostic characters highlighted are also available from <https://osf.io/zy38s/>.
Family Papilionidae Latreille, [1802]
Parnassius smintheus behrii W. H. Edwards, 1870, revised status
Genomic sequencing of Parnassius phoebus (Fabricius, 1793) (type locality in Russia: Altai) and relatives reveals that Parnassius behrii W. H. Edwards, 1870 (type locality USA: California, Tioga Pass) is placed among subspecies of Parnassius smintheus E. Doubleday, 1847 (type locality Canada: Alberta, nr. Rock Lake), rendering P. smintheus paraphyletic (Fig. 1, the Z chromosome tree). Moreover, P. behrii is not strongly differentiated genetically from various subspecies of P. smintheus. E.g., COI barcodes of P. behrii neotype (NVG-20125E02) and P. smintheus smintheus from Canada: Alberta (NVG-19083G08) differ by 0.6% (4 bp). We see that the two northwestern subspecies of P. smintheus i.e., P. s. sternitzkyi McDunnough, 1937 (type locality in USA: California, Siskiyou Co.) and P. s. olympianna Burdick, 1941 (type locality in USA: Washington, Clallam Co.), are more differentiated from the nominotypical subspecies in nuclear DNA than the nominotypical P. smintheus from P. behrii (Fig. 1). Therefore, we propose to treat P. behrii as a subspecies of P. smintheus: Parnassius smintheus behrii W. H. Edwards, 1870, stat. rev., which appears to be a more isolated geographically and genetically bottlenecked (i.e., comparatively longer branch leading to the last common ancestor of sequenced P. s. behrii specimens in Fig. 1) group of populations rather than a reproductively isolated species.
Fig. 1.

Parnassius phoebus (blue, above) and Parnassius smintheus (red and magenta, below) including Parnassius smintheus behrii (magenta).
Furthermore, barcodes of P. smintheus (NVG-19083G08) and Parnassius phoebus (Fabricius, 1793) male “neotype” in MFNB (NVG-21128G07) differ by 2.1% (14 bp), which is consistent with them being distinct species in the presence of phenotypic differences. Finally, the status of Parnassius sacerdos Stichel, 1906 (type locality in the Alps) as a species also appears questionable (Fig. 1), and it is possible that P. sacerdos may be a subspecies of P. phoebus as traditionally treated. The specimens from Switzerland (PAOE12) and Altai (NVG-21128G07) exhibit COI barcode difference of 0.9% (6 bp), which drops to 0.3% (2 bp) between P. sacerdos (PAOE12) and the lectotype of Parnassius phoebus golovinus W. Holland, 1930 (type locality USA: Alaska, Golovin Bay, NVG-21018C12) and appears to represent individual variation in mitochondrial genome rather than to stem from reproductive isolation. Genomic sequencing of larger sample of specimens throughout the ranges of these taxa is needed to confidently address these questions.
Family Pieridae Swainson, 1820
Glutophrissa drusilla noroesta Grishin, new subspecies
http://zoobank.org/3F7989B4-DB04-402D-9D82-1DAAAC62F377
Fig. 2.

Glutophrissa drusilla noroesta (red) among relatives labeled in different colors: G. d. neumoegenii (green), specimens from Suriname (magenta), and Peru (purple).
Fig. 3.

Glutophrissa drusilla noroesta ssp. n. holotype ♂, dorsal (left) and ventral (right) views, NVG-17116G07, data in text.
Fig. 4.

Glutophrissa drusilla noroesta ssp. n. ♀ from USA: AZ, Cochise Co., Hereford. iNaturalist observation 7516603. © Bob Behrstock, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Definition and diagnosis. Genomic sequencing of Glutophrissa drusilla (Cramer, 1777) (type locality likely in Suriname) specimens across the range reveals that those from Texas and Mexico form a separate clade in the Z chromosome tree (Fig. 2) sister to eastern US (Glutophrissa drusilla neumoegenii (Skinner, 1894), type locality USA: Florida, Indian River) and Caribbean Islands subspecies, rather than grouping with South American specimens that include similar in appearance Glutophrissa drusilla tenuis (Lamas, 1981) (type locality in Peru), the name currently applied to these northwesternmost populations. Therefore, the northwestern populations are not G. d. tenuis (they are not monophyletic with it) and, because no available name applies to them, are a new subspecies defined by its own clade in the Z chromosome tree. Typical males are spotless (Fig. 3), similar to eastern US and Caribbean subspecies, without an area covered in black scales by forewing apex characteristic of G. d. tenuis (Lamas, 1981) or more extensive patch of most South American populations, but with narrowly brown costal margin and outer margin from apex to about and at times a little beyond vein CuA1, frequently missing in eastern populations. More extensively colored females develop hindwing marginal border unlike eastern subspecies, and discal forewing cell rarely dark, if so, then only in part (Fig. 4). Due to extensive individual and seasonal variation, best identified by DNA sequences with the following characters in the nuclear genome: pra547.4.1:G270A, pra54.2.5:A63G, pra4.54.3:A63G, pra547.4.2: A276G, and pra828.50.5:G57A. COI barcodes do not distinguish this subspecies from others.
Barcode sequence of the holotype: Sample NVG-17116G07, GenBank OP381659, 658 base pairs: AACTCTTTATTTTATTTTTGGAATTTGATCTGGAATAGTAGGAACATCTCTAAGTTTATTAATTCGAACAGAATTAGGAAACCCTGGATCTTTAATTGGAGATGATCAAATTTATAATACTATTGTTACTGCTCATGCTTTTATTATAATTTTCTTTATAGTTATACCTATTATAATTGGAGGATTTGGAAACTGATTAGTTCCTTTAATACTTGGAGCCCCTGATATAGCTTTCCCTCGAATAAATAATATAAGATTTTGATTACTTCCCCCTTCTTTAACATTATTAATTTCAAGAAGAATTGTTGAAAATGGAGCTGGAACAGGATGAACAGTTTACCCCCCACTTTCATCTAATATTGCCCATAGTGGTTCTTCTGTTGACTTAGCTATTTTTTCTTTACACTTAGCTGGAATTTCATCAATTTTAGGAGCTATTAATTTTATTACTACTATTATTAATATACGAATTAATAATATATCATTTGATCAAATACCTCTTTTTGTTTGAGCTGTTGGTATTACTGCTCTTCTTCTTCTTCTTTCTTTACCAGTATTAGCTGGAGCTATTACTATATTATTAACTGATCGAAATTTAAATACTTCCTTCTTTGATCCCGCTGGAGGAGGAGATCCTATTTTATACCAACATTTATTT
Type material. Holotype: ♂ deposited in the Texas A&M University Insect Collection, College Station, Texas, USA (TAMU), bears five rectangular printed labels, four white: [ TEXAS: | Cameron County | Brownsville ], [ coll. | 18 Oct 1976 | Roy O. Kendall | & C. A. Kendall ], [ PIERIDAE: Pierinae | Appias drusilla neumoegeni | (Skinner, 1894) | det. Roy O. Kendall | M. & B. No. 331b ], and [ DNA sample ID: | NVG-17116G07 | c/o Nick V. Grishin ], and one red [ HOLOTYPE ♂ | Glutophrissa drusilla | noroesta Grishin ]. Paratypes: 1♀ NVG-17116G06, USA: Texas, Bexar Co., San Antonio, 31-Oct-1996, R. O. Kendall [TAMU] and Mexico: 1♂ NVG-21058B10 Sonora, Aduana, 21-Aug-1997, J. P. Brock and 1♀ NVG-21058B11 Sinaloa, Mazatlán, 2-Jan-1974, J. P. Brock.
Type locality. USA: Texas, Cameron County, Brownsville.
Etymology. This subspecies occupies the northwestern part of the species range, hence the name, which is a feminine adjective formed from “noroeste” for “northwest” in Spanish and Portuguese.
Distribution. USA: Texas through Mexico and Central America.
Neotype designation for Mylothris margarita Hübner, [1825]
Out of available names, only one, Mylothris margarita Hübner, [1825], was published without specifying type locality (Hübner [1825]), which still remains undefined. We searched for M. margarita syntypes in the collections of the Muséum National d’Histoire Naturelle, Paris, France, the Natural History Museum, London, and the Museum für Naturkunde, Berlin, Germany, but none were found, and we believe they were lost, together with most other type material of Hübner names (Hemming 1937; Calhoun 2018). Therefore, we proceeded with the neotype designation, because there is an exceptional need to define M. margarita objectively: a new subspecies proposed above and others are similar to M. margarita, and, without the type locality defined for it, potential for destabilization of nomenclature exists. According to its original illustrations, M. margarita is mostly white with dark forewing apex, some dark overscaling at forewing base by costal margin, pale-yellowish ventral forewing, and orange humeral area of ventral hindwing. It is currently regarded as a junior subjective synonym of Glutophrissa drusilla (Cramer, 1777) (type locality likely in Suriname), together with very similar to it Pieris ilaire Godart, 1819 (type locality in Brazil). To stabilize this treatment, N.V.G. designates the lectotype of Pieris ilaire Godart, 1819 as the neotype of Mylothris margarita Hübner, [1825]. As a result, the type locality of M. margarita is in Brazil, and the latter name becomes a junior objective synonym of the former.
Our neotype of M. margarita satisfies all requirements set forth by the ICZN Article 75.3, namely: 75.3.1. It is designated to clarify the taxonomic identity of Mylothris margarita Hübner, [1825], which has been in question due to similarities of the original illustrations with other named taxa in this complex, and to define its type locality that was not specified when the name was proposed; 75.3.2. The characters for the taxon include white wings with dark forewing apex and orange humeral area of ventral hindwing; 75.3.3. The neotype specimen is a male bearing four labels [ SYN- | TYPE ], [ TYPE ], [ MUSEUM PARIS | Brésil ], and [ Ilaire Goda | Bresil ]; 75.3.4. Our search for syntypes is described above, it was unsuccessful, and therefore we believe that they were lost; 75.3.5. The neotype is consistent with the original drawings in the characters given above and differs only in less yellow ventral forewing; 75.3.6. The neotype is from Brazil, which becomes the type locality of M. margarita. The type locality was not specified when the name was proposed and remained unknown; 75.3.7. The neotype is in the collection of the Muséum National d’Histoire Naturelle, Paris, France (MNHP).
Pieris marginalis siblanca Grishin, new subspecies
http://zoobank.org/97C700CF-A822-4A7F-A12C-1DAF12F30D57
Fig. 5.

Pieris marginalis marginalis (magenta) with its southeastern subspecies P. m. ziegleri with P. m. pallidissima (olive), P. m. mogollon (blue), P. m. macdunnoughii (green), and P. m. siblanca ssp. n. (red).
Fig. 6.

Pieris marginalis siblanca ssp. n. holotype, dorsal (left) and ventral (right) views, NVG-15116C06, data in text.
Fig. 7.

The type series of Pieris marginalis siblanca ssp. n. from USA: New Mexico, Sierra Blanca and Sacramento Mountains (Lincoln and Otero Cos). The holotype is shown in g and g’, others are paratypes. a. NVG-20102H01; b. NVG-20102G08; c. NVG-20102G12; d. NVG-20102G09; e. NVG-20102G10; f. NVG-20102G11; g. NVG-15116C06; h. NVG-20102H02; a–e are males and f–g are females, dorsal (left image) and ventral (right image, labels with prime, e.g., a’) views; data in text. Specimens were photographed together as a single image on a slightly greenish background, not assembled in Photoshop.
Definition and diagnosis. Sequencing of Pieris marginalis Scudder, 1861 (type locality in USA: Washington, Jefferson Co.) specimens across the range reveals genetic distinction of the population from the Sacramento mountains (Fig. 5 red), that is more different from Pieris marginalis mogollon Burdick, 1942 (type locality USA: New Mexico, Catron Co., Mogollon Range) (Fig. 5 blue) than P. m. mogollon from Pieris marginalis macdunnoughii C. Remington, 1954 (type locality USA: Colorado, San Juan Co. Silverton) (Fig. 5 green) or from Pieris marginalis pallidissima W. Barnes & McDunnough, 1916 (type locality USA: Utah, Provo) (Fig. 5 olive). Currently, the Sacramento Mts. populations that according to their genomics represent a distinct taxon, are associated with P. m. mogollon and no name is available for them. Hence, these populations represent a new subspecies, because other taxa of comparable genetic differentiation are treated as subspecies of P. marginalis. Phenotypically, this new subspecies differs from others by absent or less developed dark spots, especially at the forewing apex, and most females are spotless, only veins are outlined by dark scales, both dorsally and ventrally (Figs. 6, 7), but this dark overscaling of veins is typically more extensive than even in P. m. mogollon. In typical females (Figs. 6, 7f–h), continuous apical dark area is absent, but veins are heavily overscaled with dark-brown towards the apex, both dorsally and ventrally. Males (Fig. 7a–e) are whiter than yellower females and additionally differ from females by reduced dark overscaling on the dorsal side, in particular in the discal area of dorsal forewing, but possess more extensive overscaling at the apex, which in some specimens merges into a continuous dark apical patch towards the outer margin. Because females are easier to distinguish from other populations than males, a female is chosen as the holotype. Due to individual variation, these differences are expected to be statistical, and the new subspecies can be confidently diagnosed by a combination of the following DNA characters in the COI barcode: G34A, C64T, C115T, A415G, and 634T(not C).
Barcode sequence of the holotype: Sample NVG-15116C06, GenBank OP231464, 658 base pairs: AACTTTATATTTTATCTTCGGAATTTGATCAGGAATAGTAGGAACATCTTTAAGTTTACTTATTCGAACTGAATTAGGAAATCCAGGATTTTTAATTGGTGATGACCAAATTTATAATACTATTGTAACAGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCTATTATAATTGGAGGATTTGGAAATTGATTAGTCCCATTAATACTAGGAGCTCCAGATATAGCTTTCCCCCGAATAAATAATATAAGATTTTGATTATTACCTCCTTCTTTGACTCTTCTTATTTCAAGCAGAATCGTAGAAAATGGAGCAGGAACAGGATGAACAGTGTACCCCCCACTCTCATCAAATATTGCTCATAGAGGCTCATCAGTAGATTTAGCTATTTTTTCTTTACATTTAGCTGGGATTTCTTCAATTTTAGGAGCAATTAATTTTATTACAACTATTATTAATATACGTATTAGAAATATATCTTTTGATCAAATACCATTATTTGTATGATCAGTAGGAATTACTGCTTTACTTTTACTTCTTTCTTTACCAGTACTTGCAGGTGCAATTACAATACTTTTAACAGATCGAAATTTAAATACATCATTTTTTGATCCTGCTGGAGGAGGTGATCCAATTCTTTATCAACATCTATTT
Type material. Holotype: ♀ deposited in the C. P. Gillette Museum of Arthropod Diversity, Colorado State University, Fort Collins, CO, USA (CSUC), bears five rectangular labels: four white [ 2-VI-95 10,000’ RWH & SJC | Sierra Blanca Ski Area HQ | E slope, Sacramento Mts. | Lincoln Co., NM ], [ 19180 RWH | P. napi | mogollon ], [ DNA sample ID: | NVG-15116C06 | c/o Nick V. Grishin ], [ CSU_ENT | 1049135 ], and one red [ HOLOTYPE ♀ | Pieris marginalis | siblanca Grishin ]. Paratypes: 2♂♂ and 2♀♀ from Lincoln County, Sierra Blanca Mts.: ♂ NVG-20102G12, CSU_ENT1049153, from the type locality with the same data; ♂ NVG-20102H01, CSU_ENT1049145, from the type locality, 13-May-1974; ♀ NVG-20102G11, CSU_ENT1049142, Nogal Canyon, 7000’, 5-May-1974; ♀ Philadelphia Cyn., below Bonito Lake, 6700’, 18-Apr-1974; and 3♂♂ from Otero County, Sacramento Mts.: NVG-20102G08, CSU_ENT1049157, Five Spring, 7500’, 28-Apr-1972; NVG-20102G09, CSU_ENT1049139, 1 mi E Head Springs, 7000’, 19-Apr-1974; NVG-20102G10, CSU_ENT1049158, Mescalero Apache Reservation, Head Springs, 7000’, 9-Apr-1995 in New Mexico, USA (Figs. 5, 7), all collected by Richard W. Holland (NVG-20102G10 and NVG-20102G12 together with Steve J. Cary) and are in CSUC.
Type locality. USA: New Mexico, Lincoln County, E slope of Sierra Blanca Mountains, Ski Apache Resort Headquarters, elevation 10,000’.
Etymology. The name refers to the type locality in the Sierra Blanca Mountains, to the white color of this subspecies, to the “blank” appearance without characteristic spotting present in many Pieris marginalis populations. The name is a feminine adjective.
Distribution. Sierra Blanca and Sacramento Mountains in southern New Mexico.
Comments. This new subspecies is unexpectedly different genetically from nearby populations. It is likely that the white colors with only a few elements of wing pattern present and significant variation across and within Pieris marginalis populations hindered the discovery of this taxon.
Family Lycaenidae [Leach], [1815]
Lafron Grishin, 2020 is a subgenus of Lycaena [Fabricius], 1807
Genomic analysis of Papilio orus Stoll, 1780, the type species of Lafron Grishin, 2020, reveals that it originates within the rapid radiation at the origin of the genus Lycaena [Fabricius], 1807 (type species Papilio phlaeas Linnaeus, 1761), and therefore belongs to it (Fig. 8). It forms a long branch in the phylogenetic trees, reflecting its phenotypic uniqueness, and distinctness of the COI barcode, but is a taxon of equivalent rank to subgenera within Lycaena, and therefore we propose that Lafron Grishin, 2020, stat. nov. be treated as a subgenus of Lycaena [Fabricius], 1807.
Fig. 8.

The phylogenetic classification of Lycaeninae. Z chromosome-based tree is shown as unscaled (left, branch lengths proportional to the estimated number of accepted mutations) and scaled (right, branch lengths adjusted uniformly and with the same proportion throughout the tree so that the tips are placed at the same level). Specimens are in the same order in both trees and only one set of names is shown. The subfamily is divided into eight genera: Boldenaria (cyan), Melanolycaena (orange), Heliophorus (olive), Apangea (purple), Lycaena (blue), Helleia (magenta), Tharsalea (green), and Iophanus (red). Subgenera are labeled in different shades of colors used for genera. Names of new subgenera are highlighted in orange.
Alciphronia Koçak, 1992 is a subgenus of Lycaena [Fabricius], 1807
Frequently regarded as a synonym, Alciphronia Koçak, 1992 (type species Papilio alciphron Rottemburg, 1775) stands out in genomic trees as a clade at the subgenus level (Fig. 8, see next section). Hence, we propose that Alciphronia Koçak, 1992, stat. rest. be treated as a subgenus of Lycaena [Fabricius], 1807.
Genera and subgenera of Lycaeninae [Leach], [1815]
Genomic sequencing of representative species of Lycaeninae [Leach], [1815], including the type species of all available genus group names, gives a comprehensive overview of the subfamily phylogeny (Fig. 8). The tree is similar to the one we reported previously (Zhang et al. 2020), but now includes more taxa. We take the lowest (close to the root) level of most prominent (i.e., longer compared to others nearby) branches as the genus level, and the clades supported by these prominent branches (at about the same level) are defined as genera. This approach partitions the subfamily into eight genera (Fig. 8 and see the synonymic list below). The next (closer to the leaves) level of more prominent branches is taken as the subgenus level with 12 additional subgenera defined. Two subgenera are new (Fig. 8, their names highlighted in orange) and are described below. Similar results are obtained by “slicing” the scaled tree (Fig. 8 right, green line), except the curious irregularity with Apangea illustrated in Fig. 9. Apangea clade (purple) is characterized by reduced evolutionary rate, nearly two times slower than that of Heliophorus (Figs. 8 and 9 olive, the length of “level of” lines from the root to leaves of each clade is proportional to the average evolutionary rate of the clade). Genetic differentiations within the subgenus Heliophorus and the genus Apangea are approximately equal (Fig. 9 unscaled tree, yellow and magenta shading, respectively, and “differentiation in” lines, from the base of each clade to its leaves). When the tree is proportionally scaled, i.e., each segment is stretched equally (assuming evolutionary changes were slow all the time, from the last common ancestor of Apangea with Heliophorus, to the present (=leaves)), genetic differentiation in Apangea is nearly two times larger than in Heliophorus (Fig. 9, “differentiation in” lines in “Proportional stretching” tree with yellow and magenta shading highlighted). However, if we assume that evolution in Apangea was slower only along the branch before the diversification of the genus (from the last common ancestor of Apangea with Heliophorus to the green point on the purple branch), and therefore only this branch should be stretched, the result preserves about equal genetic differentiation in Apangea and Heliophorus (Fig. 9 “differentiation in” lines in “Preserving the two differentiations” tree with yellow and magenta shading highlighted). If proportional scaling reflects evolutionary events better, then Apangea may need divisions into subgenera, because genetic differentiation within it (Fig. 9, highlighted in magenta in the trees) is twice as large as in the subgenus Heliophorus, (Fig. 9, highlighted in yellow in the trees), and is about the same as the genetic differentiation within the genus Heliophorus that also includes Heliophorus sena, placed in subgenus Nesa. However, if slower evolution was only along the basal branch of the Apangea clade, and after the genus started diversifying, evolutionary rates of Apangea and Heliophorus became approximately equal, then the diversification in Apangea and subgenus Heliophorus started at about the same time, and there is no need to divide Apangea into subgenera.
Fig. 9.

Unequal evolutionary rates create obstacles for tree scaling. The two clades of interest in the Z chr. tree are highlighted in yellow (evolving faster) and magenta (evolving slower) and marked with green dots.
We are not able to tell which scenario better corresponds to reality. However, phenotypic assessment of differentiation would likely follow the unscaled tree, because the number of accepted mutations is loosely correlated with phenotypic change. In the unscaled tree (Fig. 9 top), genetic differentiation in Apangea is approximately the same as in the subgenus Heliophorus, and visual phenotypic assessment of Apangea indeed gives an impression of a rather compact genus, not warranting definition of subgenera. Therefore, we favor the scaling approach that preserves differentiation (Fig. 9 bottom tree) and do not define subgenera in Apangea.
Fussia Grishin, new subgenus
http://zoobank.org/09D04BA7-B1F0-454A-84A9-087247757B99
Type species. Polyommatus standfussi Grum-Grshimailo, 1891.
Definition. Originates within Lycaena [Fabricius], 1807 (type species Papilio phlaeas Linnaeus, 1761) as a likely sister of Thersamolycaena Verity, 1957 (type species Papilio dispar Haworth, 1802), however, is distant from it, forming a taxon of the same rank (Fig. 8). Distinguished from its relatives by the following combination of characters: hindwing rounded, no tail, ventrally overscaled with cream scales, without dark-brown spots present on ventral forewing, but instead with pale-brown spots encircled with white (Fig. 10), dorsal hindwing typically with purple submarginal spots or band, or broadly purple with dark postdiscal spots and dark margin. In DNA, a combination of the following base pairs is diagnostic in nuclear genome: cce2291.12.2:A1548G, cce3368.6.2:A816C, cce303125.12.1:A5073G, cce9657.10.14:G7956A, cce9657.10.31:A51G, and COI barcode: G86A, T232C, T259C, A430T, G554A.
Fig. 10.

Lycaena standfussi from China: Qinghai, Gyêgu Tibetan. iNaturalist observation 89585202. © Daniel Shi, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Etymology. The name is a feminine noun in the nominative singular, formed from the type species name.
Species included. Only the type species.
Parent taxon. Genus Lycaena [Fabricius], 1807.
Pava Grishin, new subgenus
http://zoobank.org/3A950654-D595-460C-9E02-0E7B40B46B0C
Type species. Thecla panava Westwood, 1852.
Definition. Originates within Lycaena [Fabricius], 1807 (type species Papilio phlaeas Linnaeus, 1761) as a likely sister of Lafron Grishin, 2020 (type species of Papilio orus Stoll, 1780), but is distant from it, forming a taxon of the same rank (Fig. 8). Distinguished from its relatives by nearly straight contrasting white postdiscal band from costa to inner margin of gray, black-spotted ventral hindwing (spots encircled by white) between postdiscal row of black spots and submarginal dark lunules, well-developed and connected by orange lunules with submarginal row of black spots lined with brown and then cream-white towards outer margin (Fig. 11). In DNA, a combination of the following base pairs is diagnostic in nuclear genome: cce8519.3.3:C297T, cce349.2.3:A111G, cce320.8.1:G156A, cce2297.24.1: T1437C, cce3074.1.4:T202C, and COI barcode: T13C, G77A, G78A, A474G, T562C, and T595C.
Fig. 11.

Lycaena panava from India: Uttarakhand, Nainital. iNaturalist observation 67405185. © Shriram Bhakare, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Etymology. The name is a feminine noun in the nominative singular, formed from the type species name: Pa[na]va, or the first two syllables of its unavailable synonym Polyommatus pavana Kollar, [1844].
Species included. Only the type species.
Parent taxon. Genus Lycaena [Fabricius], 1807.
Lycaena pseudophlaeas (Lucas, 1866) is a species distinct from Lycaena phlaeas (Linnaeus, 1761), and Chrysophanus abbottii Holland, 1892 is its subspecies
Genomic tree reveals that the two African taxa Chrysophanus pseudophlaeas Lucas, 1866 (type locality in Abyssinia) and Chrysophanus abbottii Holland, 1892 (type locality in East Africa) currently treated as subspecies of Lycaena phlaeas (Linnaeus, 1761) (type locality in Sweden) (Fig. 12 red) form a well-differentiated clade sister to all others (Fig. 12 green). Fst/Gmin statistics between L. p. pseudophlaeas and European L. phlaeas are 0.48/0.015, and their barcodes differ by 2.4% (16 bp). Therefore, we propose that Lycaena pseudophlaeas (Lucas, 1866), stat. rest. is a species distinct from L. phlaeas, and place C. abbottii as its subspecies forming Lycaena pseudophlaeas abbottii (Holland, 1892), comb. nov. due to a smaller genetic differentiation between them: Fst/Gmin of 0.29/0.06, and COI barcode difference of only 0.3% (2 bp). Nevertheless, as revealed by Fst/Gmin, divergence in nuclear genomes between these subspecies is non-trivial, and additional studies will clarify their status.
Fig. 12.
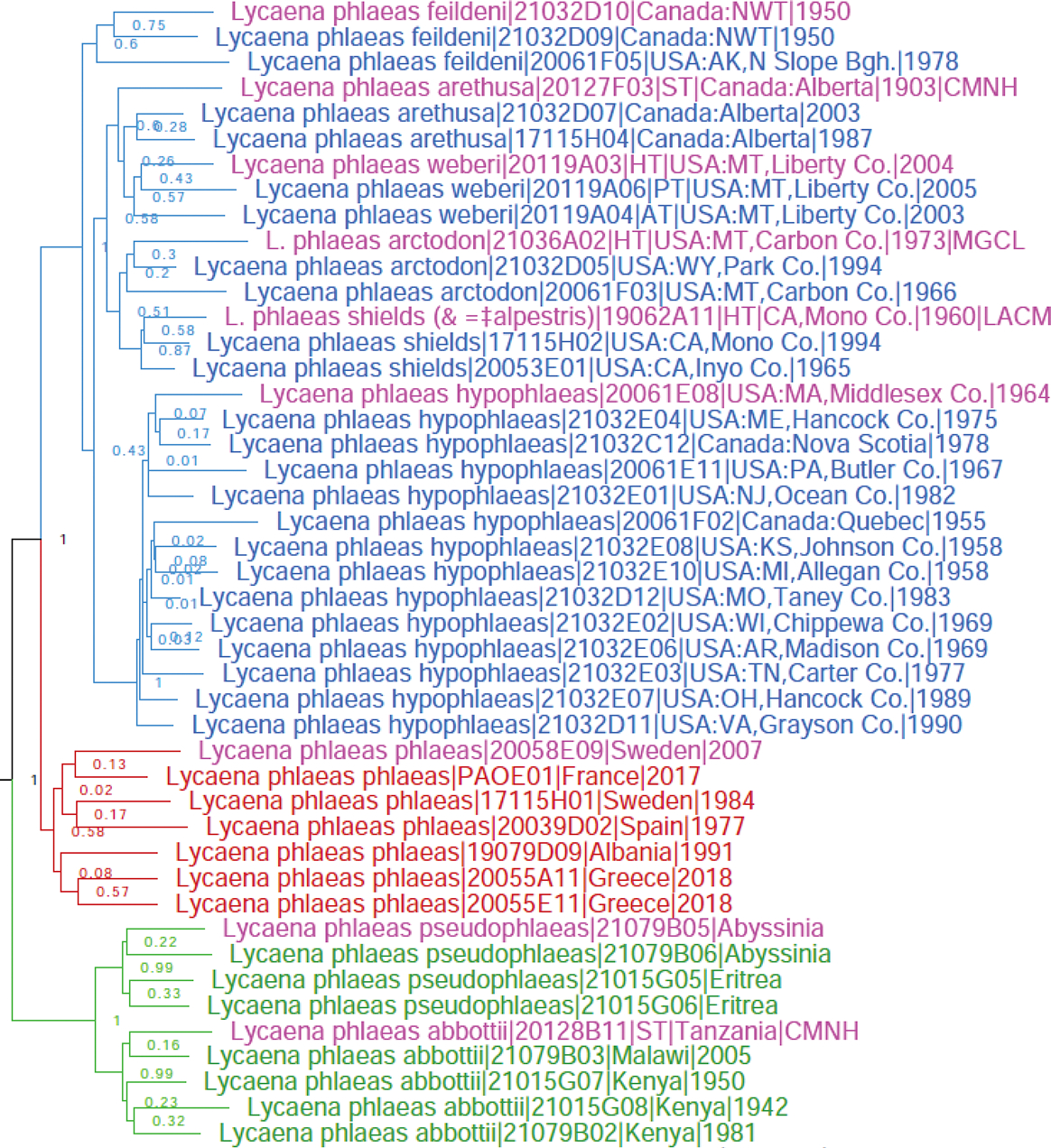
Lycaena hypophlaeas (blue), L. phlaeas (red) and L. pseudophlaeas (green). Name references are labeled in magenta.
Lycaena hypophlaeas (Boisduval, 1852) is a species distinct from Lycaena phlaeas (Linnaeus, 1761)
The Old (Fig. 12 red) and the New World (Fig. 12 blue) populations currently considered conspecific under the name Lycaena phlaeas (Linnaeus, 1761) (type locality in Sweden) form two distinct clades and are genetically differentiated with Fst/Gmin of 0.30/0.065. Although the COI barcodes differ little between them (0.8%, 5 bp), which is not unusual for species of Lycaenidae, we reinstate Lycaena hypophlaeas (Boisduval, 1852) (type locality USA: Massachusetts, vic. Boston), stat. rest., which is the oldest name for the New World populations, as a species, due to its nuclear genomic differentiation. As a result (Fig. 12), the phlaeas group consists of three species: L. phlaeas, L. hypophlaeas, and L. pseudophlaeas.
Revised Lycaeninae genera and subgenera and their available synonyms
Here, we update our previous version (Zhang et al. 2020), refining the placement of Lafron Grishin, 2020 and confirming the placement and synonymy of Phoenicurusia Verity, 1943 by genomic data. Lycaeninae are classified into eight genera and additional 12 subgenera. Changes to the previous version, except adjustment to the order of taxa, are in red font. Junior subjective synonyms are preceded by “=“, unavailable names are not listed. Type species are given in parenthesis with their original genus names.
Genus Boldenaria Zhdanko, 1995 (Lycaena boldenarum White, 1862)
Genus Melanolycaena Sibatani, 1974 (Melanolycaena altimontana Sibatani, 1974)
Genus Heliophorus Geyer, [1832] (=H. belenus Geyer, [1832], which is Polyommatus epicles Godart, [1824])
Subgenus Heliophorus Geyer, [1832] (=H. belenus Geyer, [1832], which is Polyommatus epicles Godart, [1824])
=Ilerda E. Doubleday, 1847 (Polyommatus epicles Godart, [1824])
=Kulua Zhdanko, 1995 (Polyommatus tamu Kollar, 1844)
Subgenus Nesa Zhdanko, 1995 (Polyommatus sena Kollar, 1844)
Genus Apangea Zhdanko, 1995 (Chrysophanus pang Oberthür, 1886)
Genus Lycaena [Fabricius], 1807 (Papilio phlaeas Linnaeus, 1760)
Subgenus Lafron Grishin, 2020, stat. nov. (Papilio orus Stoll, [1780])
Subgenus Pava Grishin, subgen. n. (Thecla panava Westwood, 1852)
Subgenus Fussia Grishin, subgen. n. (Polyommatus standfussi Grum-Grshimailo, 1891)
Subgenus Thersamolycaena Verity, 1957 (Papilio dispar Haworth, 1802)
Subgenus Heodes Dalman, 1816 (Papilio virgaureae Linnaeus, 1758)
=Loweia Tutt, 1906 (Papilio dorilis Hufnagel, 1766, which is Papilio tityrus Poda, 1761)
=Thersamonia Verity, 1919 (Papilio thersamon Esper, 1784)
=Palaeochrysophanus Verity, 1943 (Papilio hippothoe Linnaeus, 1760)
=Mirzakhania Koçak, 1996 (Chrysophanus kasyapa F. Moore, 1865)
Subgenus Alciphronia Koçak, 1992 (Papilio alciphron Rottemburg, 1775)
Subgenus Lycaena [Fabricius], 1807 (Papilio phlaeas Linnaeus, 1760)
Genus Helleia Verity, 1943 (Papilio helle Denis & Schiffermüller, 1775)
Genus Tharsalea Scudder, 1876 (Polyommatus arota Boisduval, 1852)
Subgenus Hyrcanana Bethune-Baker, 1914 (Polyommatus caspius Lederer, 1870)
=Sarthusia Verity, 1943 (Polyommatus sarthus Staudinger, 1866)
Subgenus Phoenicurusia Verity, 1943, confirmed status (Polyommatus phoenicurus var. margelanica Staudinger, 1881)
=Athamanthia Zhdanko, 1983, confirmed synonymy (Polyommatus athamantis Eversmann, 1854)
Subgenus Epidemia Scudder, 1876 (Polyommatus epixanthe Boisduval & Le Conte, [1835])
=Hyllolycaena L. Miller & F. Brown, 1979 (Papilio hyllus Cramer, 1775)
=Hellolycaena Koçak, 1983 (=Polyommatus thoe Guérin-Méneville, [1832], which is Papilio hyllus Cramer, 1775)
Subgenus Chalceria Scudder, 1876 (Chrysophanus rubidus Behr, 1866)
=Gaeides Scudder, 1876 (Chrysophanus dione Scudder, 1868)
Subgenus Tharsalea Scudder, 1876 (Polyommatus arota Boisduval, 1852)
Subgenus Hermelycaena L. Miller & F. Brown, 1979 (Chrysophanus hermes W. H. Edwards, 1870)
Genus Iophanus Draudt, 1920 (Chrysophanus (?) pyrrhias Godman & Salvin, 1887)
Satyrium dryope (W. H. Edwards, 1870) is a species distinct from Satyrium sylvinus (Boisduval, 1852)
Genomic tree of the subgenus Satyrium Scudder, 1876 (type species Lycaena fuliginosa W. H. Edwards, 1861) reveals that Satyrium sylvinus (Boisduval, 1852) (type locality in USA: California, Plumas Co.) may be paraphyletic with respect to Satyrium californica (W. H. Edwards, 1862) (type locality in USA: California, Napa Co.) (Fig. 13 blue), and the clade of S. sylvinus with S. californica consists of three lineages of likely equivalent taxonomic status (Fig. 13 red, blue, and green), not two. The divergence of COI barcodes among the three taxa is low, 0.5–0.6% (3–4 bp). However, it is not uncommon for closely related Lycaenidae species to have similar barcodes. For instance, barcodes of all American Celastrina Tutt, 1906 (type species Papilio argiolus Linnaeus, 1758) are identical to each other, no variation. There is little doubt that S. californica is a species distinct from S. sylvinus, and a similar level of genetic differentiation between northwestern (Fig. 13 red, includes nominal S. sylvinus) and southeastern (Fig. 13 red) groups of populations currently associated with S. sylvinus as that between S. californica and either of these groups of populations suggests that they represent two species, not one, with S. californica being the third. The oldest available name for the southeastern group of populations is Thecla dryope W. H. Edwards, 1870 (type locality in USA: California, Santa Clara Co.). Therefore, we propose it is a species-level taxon Satyrium dryope (W. H. Edwards, 1870), stat. rest. We sequenced representatives of all valid names associated with the S. sylvinus complex (Fig. 13, reference specimens labeled in magenta), and on the basis of genomic analysis assign the following taxa to S. dryope as its subspecies: Thecla putnami Hy. Edwards, 1877 (type locality USA: Utah, Mt. Nebo), Satyrium sylvinus megapallidum Austin, 1998 (type locality USA: Nevada: Elko Co., Elko), Thecla itys W. H. Edwards, 1882 (type locality USA: Arizona, Yavapai Co., Prescott), and Strymon sylvinus desertorum F. Grinnell, 1917 (type locality USA: California, Kern Co., Oak Creek). Only Satyrium sylvinus nootka M. Fisher, 1998 (type locality in Canada: British Columbia, Vancouver Island) remains a subspecies of S. sylvinus.
Fig. 13.

Satyrium sylvinus (red), Satyrium californica (blue), and Satyrium dryope (green). Reference specimens for the names are labeled in magenta.
Shijimiaeoides Beuret, 1958 is a junior subjective synonym of Glaucopsyche Scudder, 1872
The phylogenetic tree constructed from protein-coding regions of autosomes in the nuclear genome places Shijimiaeoides Beuret, 1958 (type species Lycaena barine Leech, 1893, which is a synonym or subspecies of Lycaena divina Fixsen, 1887) (Fig. 14 magenta) deep within Glaucopsyche Scudder, 1872 (type species Polyommatus lygdamus E. Doubleday, 1841) (Fig. 14 red) and near the type species of the genus and its closest relatives. COI barcodes of S. divina and G. lygdamus are only 2.4% (16 bp) different, the difference characteristic of species most closely related to each other. Therefore, we most confidently place Shijimiaeoides Beuret, 1958 as a junior subjective synonym of Glaucopsyche Scudder, 1872.
Fig. 14.

Genera Glaucopsyche (red, includes Shijimiaeoides, magenta), Turanana (blue, includes Micropsyche, olive) and Leptotes (green, includes Cyclyrius, orange).
Micropsyche Mattoni, 1981 is a junior subjective synonym of Turanana Bethune-Baker, 1916
Genomic phylogeny places monotypic genus Micropsyche Mattoni, 1981 (type species Micropsyche ariana Mattoni, 1981) as sister to Turanana Bethune-Baker, 1916 (type species Lycaena cytis Christoph, 1877) (Fig. 14 olive and blue). COI barcodes of M. ariana and Turanana cytis differ by only 3.2% (21 bp), the difference typical for closely related species. Therefore, we propose to treat Micropsyche Mattoni, 1981, syn. nov. as a junior subjective synonym of Turanana Bethune-Baker, 1916.
Cyclyrius Butler, 1897 is a junior subjective synonym of Leptotes Scudder, 1876
Despite its unusual wing patterns, Cyclyrius Butler, 1897 (type species Polyommatus webbianus Brullé, 1839) clusters closely with Leptotes Scudder, 1876 (type species Lycaena theonus Lucas, 1856, which is a subspecies of Papilio cassius Cramer, 1775) (Mérit et al. 2017), and together they form a longer branch in the tree, indicating elevated evolutionary rates compared to their relatives (Fig. 14 orange and green). COI barcodes of Cyclyrius webbianus and Leptotes cassius theonus differ by 4.6% (30 bp), which is not an uncommon difference for closely related congeners. Therefore, we confirm Cyclyrius Butler, 1897 as a junior subjective synonym of Leptotes Scudder, 1876, as it was treated by Eliot (1973).
Family Riodinidae Grote, 1895 (1827)
Apodemia cleis (W. H. Edwards, 1882), reinstated status
A genomic comparison of specimens from Arizona that includes the lectotype of Lemonias cleis W. H. Edwards, 1882 (type locality in USA: Arizona, Graham Co.) currently considered a subspecies of Apodemia zela (Butler, 1870) (type locality “Venezuela” and Mexico) with specimens from Mexico that includes a syntype of Emesis zela aureola Stichel, 1926 (type locality in Mexico: Veracruz), currently considered a junior subjective synonym of Apodemia zela zela, reveales particularly strong genetic differentiation between the two groups (Fig. 15): Fst/Gmin statistics are 0.6/0.003 and the COI barcodes between the primary type specimens differ by 6.2% (41 bp). Therefore, we reinstate the US taxon as a species Apodemia cleis (W. H. Edwards, 1882), stat. rest.
Fig. 15.

Apodemia cleis (red, above) and Apodemia zela (blue, below). Primary type specimens are labeled in corresponding colors.
Mesenopsis Godman & Salvin, 1886 is a junior subjective synonym of Xynias Hewitson, 1874
Correcting a mistake made in Zhang et al. (2021), we state that Mesenopsis Godman & Salvin, 1886 is a junior subjective synonym of Xynias Hewitson, 1874. These two names were swapped in Zhang et al. (2021), and this error is corrected here to follow the priority of the two names (1874 vs. 1886). The arguments for their synonymy are the same as presented previously (Zhang et al. 2021). We are grateful to Gerardo Lamas for kindly informing us about this error.
Family Nymphalidae Rafinesque, 1815
Aremfoxia Real, 1971 is a subgenus of Epityches D’Almeida, 1938
Genomic analysis of rarely encountered Aremfoxia Real, 1971 (type and the only species Leucothyris ferra Haensch, 1909, but see below) in the context of its relatives (Fig. 16, the tree built from Z chromosome-encoded genes) reveals close relationship with Epityches d’Almeida, 1938 (type and the only species Tritonia eupompe Geyer, 1832). COI barcodes of the Aremfoxia ferra and Epityches eupompe differ by 4.6% (30 bp), which is rather small difference even for congeners. Furthermore, provided that these closely related genera are monotypic (but see below), we feel that it is more informative for the users of taxonomic classification to reflect the close evolutionary connection between them through the common genus name, rather than keeping them in separate genera. However, taking into account phenotypic differences between these two species (A. ferra and E. eupompe), e.g., in wing shape and venation, instead of synonymizing Aremfoxia, we propose to treat it as a subgenus of Epityches.
Fig. 16.

Genera Epityches (red, includes Aremfoxia) and Pagyris (blue, includes Placidina) among others, shown in different colors. Yellow highlight illustrates genetic differentiation in some genera.
Placidina D’Almeida, 1928 is a subgenus of Pagyris Boisduval, 1870
Genomic phylogeny reveals that a monotypic genus Placidula d’Almeida, 1922 (type species Ithomia euryanassa C. Felder & R. Felder, 1860) is closely related to Pagyris Boisduval, 1870 (type species Ithomia ulla Hewitson, 1857) (Fig. 16, Z chromosome tree). COI barcodes of Placidula euryanassa differ from those of Pagyris cymothoe (Hewitson, 1855) and Pagyris ulla by 8.2% (56 bp) and 9% (59 bp), respectively. This is a moderately large difference for congeners that is more than expected from their nuclear genome differentiation (Fig. 16 blue). This differentiation between Placidula and Pagyris (Fig. 16 yellow highlight on the blue clade) is smaller than that of other related genera such as Ithomia Hübner, 1816 (type species Ithomia drymo Hübner, 1816) (Fig. 16 yellow highlight on the green clade) and Hypothyris Hübner, 1821 (type species Nerëis ninonia Hübner, [1806]) (Fig. 16 yellow highlight on the magenta clade). We believe that monotypic genera should be reserved for species that are particularly distinct from others genetically and do not have phenotypically apparent relatives, thus stressing uniqueness of such species. Therefore, we propose that Placidina D’Almeida, 1928 is a subgenus of Pagyris Boisduval, 1870. We do not consider them synonymous due to phenotypic differences, most notably more extensively scaled wings in Placidina. However, it is not unprecedented for congeners to differ in the amount of scaling, e.g., in Olyras Doubleday, 1847 (type species Olyras crathis Doubleday, 1847) and Hyalyris Boisduval, 1870 (type species Ithomia coeno Doubleday, 1847).
Epityches thyridiana (Haensch, 1909), new combination and new status
Proposed by Haensch as a form (i.e., subspecies) from Bolivia of concurrently described Leucothyris ferra Haensch, 1909 (type locality in southern Peru) and kept at this status since, thyridiana exhibits 2.4% (16 bp) difference in COI barcode from the nominotypical subspecies. Augmented with clear phenotypic differences between ferra and thyridiana mentioned by Haensch (1909) that likely stem from nuclear genome differentiation (Fig. 16), this barcode difference suggests that it is a species-level taxon Epityches thyridiana (Haensch, 1909), comb. nov., stat. nov.
Argynnis bischoffii W. H. Edwards, 1870 is a species distinct from Argynnis mormonia Boisduval, 1869
We obtained whole genome shotgun sequences of primary types of all available names currently associated with Argynnis mormonia Boisduval, 1869 (type locality probably in USA: Nevada, Washoe Co.)—except A. mormonia kimimela Marrone, Spomer & J. Scott, 2008, which is represented by a specimen within 10 miles from its type locality USA: South Dakota, Lawrence Co., Terry Peak—complemented with at least one specimen collected more recently for each name that is considered valid by Pelham (2022). A phylogenetic tree constructed from protein-coding regions of the Z chromosome reveals that Argynnis mormonia (Fig. 17 red and blue) may be paraphyletic with respect to Argynnis edwardsii Reakirt, 1866 (type locality in USA: Colorado) (Fig. 17 green), and if not (the support values are low, suggesting incomplete lineage sorting and/or gene exchange), then simply partitions this complex into three lineages of comparable genetic differentiation (Fig. 17 blue, green, and red) with Fst/Gmin statistics of 0.32–0.4/0.02–0.04. Therefore, these three lineages are likely to be species-level taxa: A. edwardsii (hardly anyone would question its distinctness), A. mormonia, and the third is Argynnis bischoffii W. H. Edwards, 1870, stat. rest. (type locality USA: Alaska, Kodiak), which is the oldest name in the blue clade (Fig. 17). Because our genetic analysis included primary types of nearly all available names, we are able to confidently assign the synonymy in the A. mormonia complex (Fig. 17) and place the following 7 taxa treated as valid by Pelham (2022) as subspecies of A. bischoffii: Argynnis opis W. H. Edwards, 1874 (type locality in Canada: British Columbia), Argynnis bischoffi [sic] washingtonia W. Barnes & McDunnough, 1913 (type locality in USA: Washington, Pierce Co.), Argynnis eurynome var. erinna W. H. Edwards, 1883 (type locality USA: Washington, Spokane Co.), Argynnis (Speyeria) mormonia kimimela Marrone, Spomer & J. Scott, 2008 (type locality in USA: South Dakota, Lawrence Co.), Argynnis eurynome W. H. Edwards, 1872 (type locality in USA: Colorado, Park Co.), Argynnis artonis W. H. Edwards, 1881 (type locality in USA: Nevada, Elko Co.), Argynnis eurynome luski W. Barnes & McDunnough, 1913 (type locality in USA: Arizona, White Mts.). Only Speyeria mormonia obsidiana J. Emmel, T. Emmel & Mattoon, 1998 (type locality in USA: California, Mono Co.) remains a subspecies of A. mormonia.
Fig. 17.

Argynnis bischoffii (blue), A. mormonia (red), and A. edwardsii (green). Specimens used as references for the names are labeled in magenta.
Argynnis leto Behr, 1862 is a species distinct from Argynnis cybele (Fabricius, 1775)
Genomic comparison strongly supports monophyly of taxa currently placed as subspecies of Argynnis cybele (Fabricius, 1775) (type locality USA: New York City), but partitions them into two distinct groups: western and eastern (Fig. 18 red and blue). Fst/Gmin statistics of 0.34/0.068 suggest species-level status of these groups. Specimens that have the appearance of “intergrades” between the two groups (Fig. 18 labeled in cyan) from the localities where the two species may meet, are confidently assigned to one of the clades and do not fall between the clades as hybrids would. Nevertheless, the “cybele leto intergrades” Passos & Grey, 1945 (type locality USA: Utah, Salt Lake City, City Creek Canyon), Speyeria cybele pugetensis F. Chermock & Frechin, 1947 (type locality USA: Washington, Mason Co.), and Speyeria cybele eileenae J. Emmel, T. Emmel & Mattoon, 1998 (type locality in USA: California: Humboldt Co.). All other subspecies considered valid by Pelham remain with A. cybele. Interestingly, New Mexican and Colorado populations are A. cybele, although they are separated from eastern populations by a larger gap in the distribution than from A. leto.
Fig. 18.

Argynnis leto (red) and Argynnis cybele (blue). Specimens used as references for the names and those that look like intergrades are labeled in magenta and cyan, respectively. A. cybele neomexicana ssp. n. is shown in olive color.
Acidalia leto valesinoides-alba Reuss, [1926] and Acidalia nokomis valesinoides-alba Reuss, [1926] are infrasubspecific names and are unavailable
Acidalia leto valesinoides-alba Reuss, [1926] and Acidalia nokomis valesinoides-alba Reuss, [1926] were proposed in the following sentence by Reuss ([1926]): “Die ♀♀ gehören zu den extremsten valesina - Formen, und benenne ich die weißen bis gelblichweißen und schwarzen ♀♀ als leto valesinoides-alba m. und nokomis valesinoides-alba m., Typen im Berliner Museum”, which we translate as: “The ♀♀ belong to the most extreme valesina - forms, and I name the white to yellowish-white and black ♀♀ as leto valesinoides-alba m[ihi] and nokomis valesinoides-alba m[ihi], types in the Berlin Museum.” No other mention of these names was made. Reuss states explicitly that he names females of a particular color variation that spans species boundaries, and names them using the same epithet (would be homonyms if available) for both species (leto and nokomis), like the name “alba” that applies to white form females of various Colias [Fabricius], 1807 species. Reuss refers to individual variation in females, not to subspecies. In the same work, Reuss ([1926]) also named a subspecies, listing it as “castetsoides n. ssp. T. Rß.”, and referred to “valesina-Formen der Weibchen” [valesina-forms of females] in contrast to that. The content of Reuss’ work unambiguously reveals that these female form names were proposed for infrasubspecific entities, and, according to the Art. 45.6.4. of ICZN Code (1999), may be unavailable. These two names were not adopted as valid for species or subspecies, and were only listed in synonymy, so Art. 45.6.4.1. does not apply. Therefore, Acidalia leto valesinoides-alba Reuss, [1926] and Acidalia nokomis valesinoides-alba Reuss, [1926] are infrasubspecific names and are unavailable.
Acidalia leto valesinoides-alba Reuss, [1926] is Argynnis leto letona (dos Passos & Grey, 1945) and not Argynnis leto leto Behr, 1862
Genomic analysis of the specimen selected as the “holotype” Acidalia (Semnopsyche) leto valesinoidesalba Reuss, [1926] (type locality not stated, NVG-18055C12 ♀ in MFNB labeled from Provo, Fig. 19) places it among specimens of Argynnis leto letona (dos Passos & Grey, 1945) (type locality USA: Utah, Salt Lake City, City Creek Canyon) (Fig. 18) in agreement with its label data and implying that it is not synonymous with Argynnis leto Behr, 1862 (type locality Nevada, nr. Carson City) as currently treated. Therefore, infrasubspecific name A. l. valesinoides-alba Reuss, [1926] should be listed among unavailable names associated with Speyeria leto letona dos Passos & Grey, 1945. The “type locality” of A. l. valesinoides-alba is USA: Utah, Utah County, Provo according to the label of its “holotype”. Quotes are used here because unavailable names do not formally have holotypes or type localities.
Fig. 19.

“Holotype” of Acidalia leto valesinoides-alba Reuss, [1926], NVG-18055C12, dorsal (left) and ventral (right) views with labels. All images are to scale except the locality label (with “Provo”) that is reduced, and its scale is shown above it.
Argynnis cybele neomexicana Grishin, new subspecies
http://zoobank.org/DBD207B7-4FC2-4A90-B42E-5ED6EC58A1E0
(Figs. 18 part, 20 part, 21, 22)
Fig. 20.
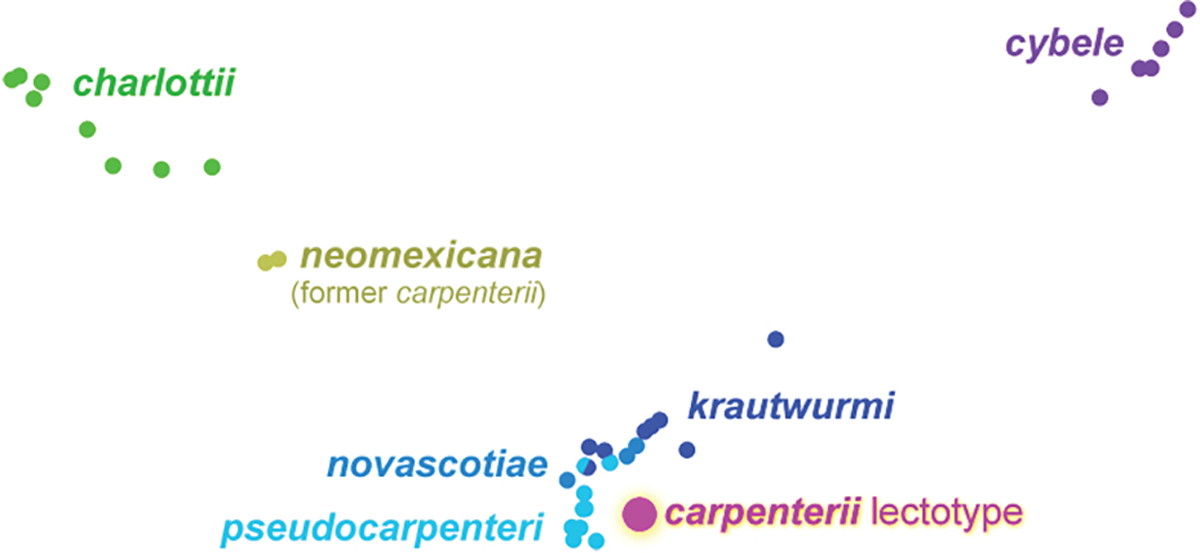
PCA analysis of Argynnis cybele: carpenterii lectotype (magenta), neomexicana ssp. n. (olive), charlottii (green), cybele (purple), and others (shades of blue), carried out as described in Cong et al. (2021).
Fig. 21.

Holotype of Argynnis cybele neomexicana ssp. n. dorsal (left) and ventral (right) views, data in text.
Fig. 22.

Argynnis cybele neomexicana ssp. n. from USA: New Mexico, Sandoval Co. iNaturalist observations 73138385 (left), 73138564 (right). © Ken Kertel, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Definition and diagnosis. Sequencing of the lectotype of Argynnis cybele carpenterii W. H. Edwards, 1876 (NVG-20126C08, labeled from “top Taos Mtn NM”) reveals that it is not grouping in the tree (Fig. 18) or PCA analysis (Fig. 20) with the specimens we sequenced from New Mexico or even Colorado and Utah, and therefore does not belong to the north-central New Mexican populations, contrary to the current understanding. Instead, the Argynnis cybele carpenterii lectotype is placed within more eastern specimens from the US and Canada, and we hypothesize that it was either mislabeled, or the Taos Peak population (which we have not sequenced) is the southernmost remnant of northeastern A. cybele. In either case, the New Mexican subspecies referred to as A. c. carpenterii (Fig. 18 olive) is left without a name, which is proposed here. This new subspecies is similar in appearance to Argynnis cybele charlottii W. Barnes, 1897 (type locality USA: Colorado, Garfield Co., Glenwood Springs) but is genetically distinct from it, and can be distinguished by typically larger silver spots on ventral hindwing (especially in males compared to typical males of A. c. charlottii), narrower cream band between postdiscal and submarginal rows of silver spots, silver spots at forewing apex beneath (usually), and less prominent dark overscaling at wing basal halves above (Figs. 21, 22).
Barcode sequence of the holotype: Sample NVG-21022E12, GenBank OP231465, 658 base pairs: GACTTTATATTTTATTTTTGGGATTTGAGCAGGAATAGTAGGAACATCATTAAGTTTATTAATTCGAACTGAATTAGGTAACCCAGGGTCACTAATTGGAGATGATCAAATTTACAATACTATTGTAACAGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCAATTATAATTGGAGGATTTGGTAACTGATTAGTCCCCCTAATATTAGGAGCTCCAGATATAGCTTTCCCCCGTATAAACAATATAAGATTTTGACTTTTACCCCCATCCTTAATTTTACTTATTTCTAGAAGAATTGTAGAAAATGGAGCAGGAACAGGATGAACAGTATACCCCCCTCTTTCTTCTAATATTGCCCATAGAGGTTCTTCAGTAGATTTAGCAATTTTCTCTTTACATTTAGCAGGAATTTCTTCTATTTTAGGAGCAATTAACTTTATTACAACAATTATTAATATACGAATTAATAGAATATCTTTTGATCAAATACCATTATTTGTGTGAGCAGTAGGAATCACAGCCTTACTTCTTTTACTATCTTTACCAGTTTTAGCAGGAGCTATTACAATACTTTTAACTGATCGTAATTTAAATACTTCTTTTTTTGACCCTGCAGGAGGAGGAGACCCTATTTTATACCAACATTTATTT
Type material. Holotype: ♂ in the C.P. Gillette Museum of Arthropod Diversity, Colorado State University, Fort Collins, CO, USA (CSUC), bears four rectangular printed labels: three white [ 9-VII-83 leg. RWH 8900’ | Dome Lookout,St.Peter’s | Dome, E. slope, Jemez | Mts., Sandoval Co., NM ], [ 14877 RWH | S. cybele | carpenteri ], [ DNA sample ID: | NVG-21022E12| c/o Nick V. Grishin ], and one red [ HOLOTYPE ♂ | Argynnis cybele | neomexicana Grishin ]. It was collected by Richard W. Holland.
Paratype: ♂ NVG-21022E11 USA: New Mexico, Sandoval Co., S slope of Jemez Mts., 4 mi down Bland Canyon from Bland, elevation 6500’, 9-Jul-1983, leg. Richard W. Holland.
Type locality. USA: New Mexico, Sandoval Co., E slope of Jemez Mts., Saint Peter’s Dome, Dome Lookout, elevation 8900’.
Etymology. The name is given for the type locality that is in New Mexico. The name is a feminine adjective.
Distribution. North-central New Mexico and southwestern Colorado.
Argynnis cybele carpenterii W. H. Edwards, 1876 is from northeastern populations
“Taxonomists should not name anything pseudo-. Nearly every one of those named recently [also in Callophrys & Colias] has become embroiled in disputes.”
The genomic analysis reveals that the lectotype of Argynnis cybele carpenterii W. H. Edwards, 1876 (type locality USA: New Mexico, Taos Co., Taos Peak, possibly mislabeled) is close to the three closely related and therefore questionably distinct subspecies: Argynnis cybele krautwurmi W. Holland, 1931 (type locality in USA: Michigan, Mackinac Co.), Argynnis cybele novascotiae McDunnough, 1935 (type locality in Canada: Nova Scotia), and Argynnis cybele pseudocarpenteri F. Chermock & R. Chermock, 1940 (type locality Canada: Manitoba, Sand Ridge), but is somewhat distant from each of them (Figs. 18, 20). Sequencing of specimens from additional localities will clarify the origins of A. c. carpenterii, which we presently regard as subspecies distinct from the other three due to genetic differences.
Boloria myrina (Cramer, 1777) is a species distinct from Boloria selene ([Denis & Schiffermüller], 1775)
Genomic sequencing of Boloria selene ([Denis & Schiffermüller], 1775) (type locality Austria: Vienna) specimens from across its range reveals two distinct clades, which correspond to the Old and the New World groups of populations (Fig. 23, Z chromosome). The high genetic differentiation between the clades, low gene exchange (Fst/Gmin of 0.48/0.01), and COI barcode difference of 3.3% (22 bp) suggest that the two clades represent two species. The oldest available name for the New World species is Boloria myrina (Cramer, 1777), stat. rest. (type locality in USA, probably southeastern New York) and all North American taxa currently attributed to B. selene become subspecies of B. myrina.
Fig. 23.

Boloria myrina (red, top), and B. selene (blue, bottom).
Melitaea sterope W. H. Edwards, 1870 is a subspecies of Chlosyne palla (Boisduval, 1852) and is not conspecific with Chlosyne acastus (W. H. Edwards, 1874)
The genomic tree constructed from specimens of Chlosyne Butler, 1870 (type species Papilio janais Drury, 1782) reveals that Melitaea sterope W. H. Edwards, 1870 (type locality in USA: Oregon, Wasco Co.) (Fig. 24 magenta) currently considered conspecific with Chlosyne acastus (W. H. Edwards, 1874) (type locality in USA: Utah, probably Utah Co.) (Fig. 24 red) is not monophyletic with it, and instead originates within Chlosyne palla (Boisduval, 1852) (type locality in USA: California, Plumas Co.) (Fig. 24 blue). This conclusion is solid, because we sequenced the primary types of all three taxa in question: M. sterope, C. palla, and C. acastus to provide the ultimate reference for these names. Therefore, Melitaea sterope W. H. Edwards, 1870 is not conspecific with Chlosyne acastus (W. H. Edwards, 1874) and is a subspecies of Chlosyne palla (Boisduval, 1852): Chlosyne palla sterope (W. H. Edwards, 1870), comb. rev., which may be a welcome development for preserving the name acastus.
Fig. 24.

Chlosyne acastus (red) and C. palla (blue and magenta) with sterope (magenta) as its subspecies. Primary types are labeled in their species’ colors.
Phyciodes jalapeno J. Scott, 1998 is a species distinct from Phyciodes phaon (W. H. Edwards, 1864)
Originally proposed and kept since as a subspecies, Phyciodes phaon jalapeno J. Scott, 1998 (type locality USA: Arizona, Maricopa Co., Mesa) is genetically distinct from the nominotypical P. phaon (W. H. Edwards, 1864) (type locality USA: Georgia, Glynn Co., San Simon Isl., neotype NVG-20129G11 sequenced) (Fig. 25): Fst/Gmin statistics are 0.37/0.04 and COI barcode difference is 1.8% (12 bp). Therefore, we propose that it is a species-level taxon Phyciodes jalapeno J. Scott, 1998, stat. nov.
Fig. 25.

Phyciodes jalapeno (red, above) and P. phaon (blue, below).
Phyciodes incognitus Gatrelle, 2004, Phyciodes orantain J. Scott, 1998, Phyciodes anasazi J. Scott, 1994 (including P. batesii apsaalooke J. Scott, 1994 as a subspecies), and Phyciodes diminutor J. Scott, 1998 are species-level taxa
Genomic comparison of the four Phyciodes Hübner, [1819] (type species Papilio cocyta Cramer, 1777) that constitute the tharos species group, which are closely related and difficult to identify: Phyciodes tharos (Drury, 1773) (type locality USA: New York City), Phyciodes cocyta (Cramer, 1777) (type locality in Canada: Nova Scotia), Phyciodes batesii (Reakirt, [1866]) (type locality USA: Virginia, Winchester) and Phyciodes pulchella (Boisduval, 1852) (type locality USA: San Francisco), reveals more complex speciation scenarios than currently recognized (Fig. 26) (Scott 1994; Scott 1998; Scott 2006; Pelham 2022). While these four taxa are indeed species according to our genomics-based criteria, four other lineages are of the same rank as these four. First, Phyciodes cocyta incognitus Gatrelle, 2004 (type locality in USA: Georgia, Union Co.) (Fig. 26 orange) is not monophyletic with P. cocyta (Fig. 26 purple), but instead is sister to P. tharos (Fig. 26 blue), and due to sympatry between P. tharos and P. c. incognitus, we reinstate the latter as a species-level taxon: Phyciodes incognitus Gatrelle, 2004, stat. rest.
Fig. 26.

Trees constructed from protein-coding regions in Z-chromosome (left) and mitogenome (right) of eight Phyciodes species in the tharos group: tharos (blue), incognitus (orange), diminutor (olive), cocyta (purple), anasazi (green), orantain (red), batesii (cyan), and pulchella (magenta). Two specimens collected together shown in Fig. 27 are highlighted in yellow.
Second, Phyciodes tharos orantain J. Scott, 1998 (type locality USA: Colorado, Adams Co., Barr Lake) (Fig. 26 red) is not monophyletic with P. tharos (Fig. 26 blue) and is quite distant from all other taxa. A curious observation is that one of the specimens labeled as a paratype of P. t. orantain (NVG-21067C12, Fig. 27 top) collected at its type locality was placed within P. tharos in the tree (Fig. 26, highlighted yellow within blue clade). Puzzled about this placement, we inspected photographs of the specimen and found that it had black antenna nudum and not orange as in the namesake P. t. orantain (Fig. 26 red). Another P. t. orantain paratype with the same locality and date (NVG-21067C11, Fig. 27 bottom), but with orange nudum was placed within all other P. t. orantain specimens (Fig. 26 highlighted yellow within red clade). Thus, we demonstrate by genomic sequencing that P. t. orantain is sympatric and synchronic with P. tharos (unless that specimen was mislabeled, which is not likely because its mitogenome is that of P. t. orantain, Fig. 26 right) and here propose that it is a species-level taxon: Phyciodes orantain J. Scott, 1998 stat. nov. The paratypes illustrated in Fig. 27 are labeled in Gatrelle’s hand, so it is possible that Scott simply didn’t notice the black antenna nudum when giving these specimens to Gatrelle as “paratypes,” thus missing the opportunity to exclude this P. tharos specimen from the type series of P. orantain, or some other mishap occurred, like specimen mislabeling.
Fig. 27.

Paratypes of Phyciodes orantain with their labels: NVG-21067C12, which is Phyciodes tharos (top, black nudum); and NVG-21067C11 (bottom, orange nudum). All images are to scale, except insets showing enlarged view of antenna club.
Third, sisters Phyciodes batesii apsaalooke J. Scott, 1994 (type locality in USA: Wyoming, Bighorn Co.) and Phyciodes batesii anasazi J. Scott, 1994 (type locality in USA: Colorado, Mesa Co.) (Fig. 26 green) are not monophyletic with P. batesii (Fig. 26 cyan), but instead form a clade sister to P. cocyta (Fig. 26 purple). Due to the genetic and morphological distinction of P. b. apsaalooke with P. b. anasazi from P. cocyta, we propose to treat them as a distinct species Phyciodes anasazi J. Scott, 1994 stat. nov., with Phyciodes batesii apsaalooke J. Scott, 1994, comb. nov. as its subspecies. Here, acting as the first reviser, we gave priority to the name anasazi over apsaalooke, because the name is shorter, and the taxon has a wider distribution. The decision to elevate P. anasazi to the species level is largely prompted by its apparent phenotypic similarity with P. batesii, rather than with a closer relative P. cocyta. Genetic distinction of the former two similar species suggests hybrid origin of at least some species.
Fourth, Phyciodes cocyta diminutor J. Scott, 1998 (type locality in USA: Minnesota, Freeborn Co.) (Fig. 26 olive) is sister to the clade consisting of P. cocyta and P. anasazi, and, therefore, we confirm it as a species-level taxon: Phyciodes diminutor J. Scott, 1998. An alternative treatment may be to consider P. cocyta, P. anasazi, and P. diminutor conspecific because they are closest to each other genetically. However, each of the three species forms a distinct clade in the tree, and future studies will address the complexities of their evolution and speciation.
We observe that Z chromosome proteins are quite similar between P. batesii and P. pulchella (Fig. 26 cyan and magenta), and the two species are sisters in the Z chromosome tree. Autosomal proteins (not shown) separate these two species better, placing P. pulchella as sister to all other Phyciodes of the tharos group, which seems more in agreement with their phenotypes. Thus, the evolutionary history of the tharos group is riddled with irregularities such as hybridization and introgression.
Finally, the mitochondrial genome tree (Fig. 26 right) has an appearance of a partly scrambled version of the nuclear Z chromosome tree (Fig. 26 left), but to the extent that on the current sample of specimens it is nearly impossible to assign ancestral haplotypes to all species, although major clades of P. tharos, P. orantain, P. anasazi, and P. pulchella probably correspond to such. While it reflects to some extent the relationships observed in the nuclear genome, mitogenome cannot be used with confidence in taxonomic work and specimen identification due to extensive introgression. Apparently, the tharos group species are incipient, and they hybridize with a certain frequency, despite being mostly distinct.
Type locality of Junonia pacoma Grishin, 2020 is in Sinaloa, not Sonora
The type locality of Junonia pacoma Grishin, 2020 was incorrectly given in the text of the original description as “Mexico: Sonora, Isla de la Piedra” (Cong et al. 2020), which is here corrected to “Mexico: Sinaloa, Isla de la Piedra”. Furthermore, all references to “Sonora” in the description and illustrations of J. pacoma in that work (Cong et al. 2020) are corrected to “Sinaloa”. We are grateful to Andrew D. Warren for kindly informing us about this mistake. These localities were listed correctly in the Supporting Information Table S1 (Cong et al. 2020). Junonia pacoma has also been recorded and genetically confirmed from Sonora, e.g., a pair in UCDC collected by R. E. Wells south of San Carlos on beach dunes: NVG-19065E05 ♀ 26-Mar-2003 and NVG-19065E06 ♂ 5-Feb-2005.
Cercyonis incognita J. Emmel, T. Emmel & Mattoon, 2012 is a subspecies of Cercyonis silvestris (W. H. Edwards, 1861), reinstated status
We find that the lectotype of Cercyonis sthenele silvestris (W. H. Edwards, 1861) (type locality USA: California, suggested to be in Butte Co.) (Figs. 28 magenta, 29) is not conspecific with Cercyonis sthenele (Boisduval, 1852) (type locality USA: California, San Francisco, paralectotype sequenced) (Fig. 28 green), but is in the same clade with Cercyonis oetus (Boisduval, 1869) (type locality in USA: California, Placer Co.) and Cercyonis incognita J. Emmel, T. Emmel & Mattoon, 2012 (type locality in USA: California, Mendocino Co.) (Fig. 28 blue and red). Even the Z chromosome-based tree (Fig. 28) did not reveal prominent genetic differentiation between C. incognita and C. oetus, suggesting recent divergence of these species and posing questions about their reproductive isolation to be addressed in future work. We refrain from treating these taxa as conspecific due to the difficulty in rearing adults from crosses between them (Emmel et al. 2012). The phylogenetic analysis we performed differentiates species diverged farther back in time than C. oetus and C. incognita and may not be able to handle recently diverged species. Regardless of the status of C. oetus and C. incognita, the lectotype of C. s. silvestris (Fig. 28 magenta), together with a more recently collected specimen identified by facies as a possible C. s. silvestris (NVG-21095D08 USA: CA, El Dorado Co.), are placed in the tree with C. incognita (Fig. 28 red), and therefore could be conspecific with it. Because of wing pattern differences between C. s. silvestris (darker) and C. incognita (paler), we consider the latter to be a subspecies of the former, rather than its synonym: Cercyonis silvestris incognita J. Emmel, T. Emmel & Mattoon, 2012, stat. nov. implying that Cercyonis silvestris (W. H. Edwards, 1861), stat. rest. is reinstated here as a species. Finally, we note that C. oetus is not monophyletic in the Z chromosome tree (Fig. 28), and its nominotypical specimens are in the same clade with C. silvestris. Therefore, if C. silvestris is a species distinct from C. oetus, it is conceivable that C. oetus consists of several species.
Fig. 28.

Cercyonis silvestris (red, lectotype in magenta, incognita as subspecies), oetus (blue), sthenele (green), and meadii (purple).
Fig. 29.

Lectotype of Satyrs silvestris W. H. Edwards, 1861, NVG-20126E02, dorsal (left) and ventral (right) views, and its labels. Labels are reduced relatively to the specimen: larger and smaller scale bars refer to specimen and labels, respectively.
Paramacera allyni L. Miller, 1972 and Paramacera rubrosuffusa L. Miller, 1972 are species distinct from Paramacera xicaque (Reakirt, [1867])
Nuclear genomic tree partitions sequenced Paramacera A. Butler, 1868 (type species Neonympha xicaque Reakirt, [1867]) specimens into three distinct clades corresponding to three named taxa (Fig. 30). Here, we argue for species-level status of the three taxa: not only of Paramacera allyni L. Miller, 1972, stat. rest. (type locality USA: Arizona, Cochise Co., Barfoot Park), but also of Paramacera rubrosuffusa L. Miller, 1972, stat. nov. (type locality in Mexico: Oaxaca) initially proposed as a subspecies. COI barcodes of P. allyni and P. rubrosuffusa differ from Paramacera xicaque (Reakirt, [1867]) (type locality in Mexico: Veracruz) by 3.2% (21 bp) and 3.8% (25 bp), respectively. This level of barcode divergence (>3%), in the presence of phenotypic differences (which are apparent in Paramacera), has been suggested as sufficient to substantiate even allopatric taxa as distinct species (Lukhtanov et al. 2016). Therefore, we propose to treat the three Paramacera taxa as species.
Fig. 30.

Paramacera allyni (red), P. xicaque (blue), and P. rubrosuffusa (magenta). Primary types are labeled in corresponding colors.
Five Cissia rubricata group species, not subspecies, including a new one
Sequencing a sample of specimens representing all five available names currently referring to subspecies of Cissia rubricata (W. H. Edwards, 1871) (type locality USA: TX, McLennan Co., nr. Waco) that includes the primary types of all five names reveals prominent genetic differentiation among four of these taxa (Fig. 31), which in addition to the nominotypical are: Euptychia rubricata cheneyorum R. Chermock, 1949 (type locality USA: AZ: Pima Co., Madera Canyon), Megisto rubricata pseudocleophes L. Miller, 1976 (type locality in Mexico: Guerrero), and Megisto rubricata anabelae L. Miller, 1976 (type locality in Mexico: Chiapas). The COI barcode difference exceeds 6% for the closest pair of these taxa. Provided phenotypic distinction, this barcode difference supported by consistent clustering in the nuclear genome tree (Fig. 31 left) argue for the species, rather than subspecies, level of these taxa. Conversely, COI difference between the holotype of C. rubricata smithorum (Wind, 1946) (type locality in USA: Texas, Marfa-Alpine, NVG-15105E05) and the neotype of C. rubricata rubricata (NVG-20125F01) is 0.8% (5 bp), which, given large barcode divergence among species in the rubricata group, is comparatively small. Therefore, we keep these two taxa as subspecies, but elevate others to species: Cissia cheneyorum (R. Chermock, 1949), stat. nov., Cissia pseudocleophes (L. Miller, 1976), stat. nov., and Cissia anabelae (L. Miller, 1976), stat. nov. Moreover, we find the fifth species-level clade in the tree that was not associated with any name (Fig. 31, red) and is therefore new. It is described below as a species.
Fig. 31.

Trees constructed from protein-coding regions in Z-chromosome (left) and mitogenome (right) of five Cissia species: rubricata (green and olive) with smithorum as its subspecies (olive), cheneyorum (purple), anabelae (blue), wahala sp. n. (red), and pseudocleophes (cyan). Specimens used as references for the names are labeled in magenta.
Cissia wahala Grishin, new species
http://zoobank.org/83981016-4965-4097-A3F5-4D8C37824DFF
Fig. 32.

Holotype of Cissia wahala sp. n. dorsal (left) and ventral (right) views, NVG-14105D03, data in text.
Fig. 33.

Cissia wahala from Mexico: Puebla, Ocoyucan. iNaturalist observation 57699556. © Bodo Nuñez Oberg, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Definition and diagnosis. Considered by Miller (1976) within his concept of C. anabelae, but genetically distinct from it and other rubricata group taxa at the species level. COI barcode sequence of the holotype (NVG-14105D03) differs by 5.8% (38 bp), 6.2% (41 bp), 8.5% (56 bp), 5.9% (39 bp), and 6.7% (44 bp) from the holotype of C. anabelae, topotype of C. cheneyorum (NVG-21027E09), the holotypes of C. pseudocleophes (NVG-21042F03) and C. rubricata smithorum (NVG-15105E05), and the neotype of C. rubricata rubricata (NVG-21096C03), respectively. Distinguished from its relatives by a combination of the following characters: red patches on dorsal side of wings larger than in other species with developed mottling on ventral hindwing: e.g., forewing patch occupies more than distal posterior quarter of the forewing discal cell and extends into cells distad of it, at least partly; small or absent eyespots on hindwing below, mostly without silver pupils; mottled ventral hindwing as in C. anabelae; and colder, redder and paler ventral surface compared to warmer, yellower and darker one in C. anabelae. In COI barcode, diagnosed by a combination of the following base pairs: A49T, C268T, A382G, T484C, T581C, and A622C.
Barcode sequence of the holotype: Sample NVG-14105D03, GenBank OP231466, 658 base pairs: AACTTTATATTTTATTTTCGGAATTTGAGCAGGTATAGTAGGTACATCTCTTAGTTTAATTATTCGAATAGAATTAGGAAACCCCGGATTTTTAATTGGAGATGATCAAATTTATAATACTATTGTTACTGCTCACGCTTTTATTATAATTTTTTTTATAGTAATACCTATTATAATTGGAGGATTCGGAAACTGACTAGTCCCCTTAATACTAGGAGCCCCTGATATAGCTTTCCCCCGTATAAATAATATAAGATTTTGATTACTTCCCCCATCTTTAATTTTATTGATTTCAAGAAGTATCGTAGAAAATGGAGCTGGAACAGGATGAACTGTTTATCCCCCCCTTTCATCTAATATTGCCCATAGAGGATCCTCTGTGGATTTAGCTATTTTCTCCCTTCATTTAGCTGGAATTTCCTCAATTTTAGGAGCTATTAATTTTATTACAACAATTATTAATATACGAATTAATAGCATATCCTATGATCAAATACCCCTATTTGTCTGAGCTGTAGGGATCACAGCTCTCTTACTTCTTCTTTCTTTACCTGTTTTAGCTGGAGCAATTACTATACTTCTAACAGACCGAAATTTAAATACATCATTTTTTGACCCTGCCGGAGGAGGAGATCCAATCTTATATCAACATTTATTT
Type material. Holotype: ♂ deposited in the National Museum of Natural History, Smithsonian Institution, Washington, DC, USA (USNM), bears two white rectangular labels [ MEXICO: Oax., 6miN | Huajuapan de Leon | 6 September 1972 | G.F. & S. Hevel ] and [ DNA sample ID: | NVG-14105D03 | c/o Nick V. Grishin ] and one red [ HOLOTYPE ♂ | Cissia wahala | Grishin ]. Paratypes: 5♀♀ from Mexico, 3 from Oaxaca, around Huajuapan de Leon (NVG-21073A08, 5-Sep-1971, also a paratype of C. anabelae; NVG-1924, elev. 5900’, 10-Aug-1967, J. E. Hafernik) and 5 mi S of Matatlan on MX190 (NVG-21073B01, 24-Aug-1956, M. G. Douglas), and 2 from Puebla (NVG-14105C12, ca. 24 mi SE of Acatlan, nr. Chila, 14-Aug-1972, C F. & S. Hevel; and NVG-1922, 6.4 mi E of Azumbilla, 15-Apr-1979, T. P. Friedlander & J. C. Schaffner). Only sequenced specimens are included as paratypes.
Type locality. Mexico: Oaxaca, 6 mi north of Huajuapan de Leon.
Etymology. The name is a phonetic fusion of Oaxaca with Puebla: waha(i.e., Oaxa)[ca+Pueb]la, and it also stands for the difficulty of discovering this species and troubles in separating it from C. anabelae. The name is a feminine noun in apposition.
Distribution. Currently known only from the states of Puebla and Oaxaca in Mexico.
Family Hesperiidae Latreille, 1809
Tarsoctenus gaudialis (Hewitson, 1876) is a species distinct from Tarsoctenus corytus (Cramer, 1777)
Genomic analysis of Erycides gaudialis Hewitson, 1876 (type locality Panama: Chiriqui), currently a subspecies of Tarsoctenus corytus (Cramer, 1777) (type locality in Suriname) reveals a deep split between them (Fig. 34, the Z chromosome tree) with Fst/Gmin statistics of 0.55/0.006 and COI barcode difference between a syntype of Erycides gaudialis in MFNB (NVG-15029C08) and a specimen of nominotypical T. corytus from French Guiana (NVG-18086G01) of 4.6% (30 bp). Therefore, we reinstate Tarsoctenus gaudialis (Hewitson, 1876), stat. rest. as a species-level taxon.
Fig. 34.

Tarsoctenus gaudialis (red, above) and Tarsoctenus corytus (blue, below). Primary types are labeled in corresponding colors.
Genetic uniformity of Spicauda simplicius (Stoll, 1790) across its range
Inspection of genomic trees frequently reveals prominent genetic splits (especially in the Z chromosome) between geographically separated populations, particularly along major suture zones. We interpret these splits as speciation events and consider genetically differentiated groups of populations with limited gene exchange as species-level taxa. Examples can be found throughout this work, such as the section just above about Tarsoctenus gaudialis versus T. corytus. Here, we show an example where we were not able to find any splits or breaks in genetic differentiation across the entire range of a species. Figure 35 shows the Z chromosome tree of 28 specimens of Spicauda simplicius (Stoll, 1790) (type locality in Suriname) from USA: Texas through North, Central, and South America to Bolivia and South Brazil, including a specimen from the type locality (NVG-19121E01), that, in the absence of primary type specimens which are likely lost, is used here as a reference for this name. This vast distribution crosses several major suture zones known to separate many Hesperiidae species. However, the S. simplicius tree has an appearance of a comb and does not reveal any meaningful bifurcations, indicating the lack of genetic differentiation into discrete groups of populations that may be treated as several distinct species. Therefore, for the lack of evidence to the contrary, all these populations identified as S. simplicius represent a single species distributed from South Texas (as a stray) to Brazil. Furthermore, sequencing of two syntypes of Goniurus pilatus Plötz, 1881 (no. 5068 and 5069, type locality in Bahia, [Brazil] and Suriname) (Fig. 35) confirms that this name is a junior subjective synonym of S. simplicius. Thus, not every morphospecies would be dividable into several species after genomic analysis using our criteria of genetic differentiation and gene exchange.
Fig. 35.

Spicauda simplicius (blue) among other members of the genus. Primary type specimens are labeled in blue.
Epargyreus in northwestern North America
As we previously found (Zhang et al. 2020), resident Epargyreus Hübner, [1819] (type species Papilio tityrus Fabricius, 1775, a junior homonym, valid name for this species is Papilio clarus Cramer, 1775) is represented in the USA by two species: Epargyreus clarus (Cramer, 1775) (type locality “Suriname”, later corrected to USA: Virginia, Rockingham Co.) and Epargyreus huachuca Dixon, 1955 (type locality in USA: Arizona, Cochise Co.). Here, we clarify the status of Epargyreus populations in northwestern North America. We observed (Zhang et al. 2020) that genetic differentiation within Epargyreus clarus californicus MacNeill, 1975 (type locality USA: CA, El Dorado Co., China Flat) was substantially lower than that of the nominotypical Epargyreus clarus, suggesting a recent bottleneck and possible recolonization of the vast range of E. c. californicus (Fig. 37, red vs. blue, Z chromosome tree). To probe older distribution of these taxa in the northwestern USA, we sequenced two specimens (in CMNH) collected more than a century ago in the states of Oregon and Washington (Fig. 36). Consistent with their phenotypes discussed by Warren (2005), one was E. clarus clarus (Fig. 37 blue, NVG-21014C06) and the other was E. clarus californicus (Fig. 37 red, NVG-21014C07), supporting the hypothesis that eastern E. c. clarus reaches Oregon, and Californian E. c. californicus reaches Washington, and they did so a century ago. We also sequenced a piece of exuviae (no specimen) from pupation (on 29 July 2018) of a larva reared from an egg found on giant vetch (Vicia nigricans var. gigantea (Hook.) Broich) by Christian Gronau in Canada: British Columbia, Cortes Island, Manson’s Landing, and it was E. clarus californicus (Fig. 37, red, NVG-22021G04). The Cortes Island population (Fig. 38) extends the range of sub-species californicus, which ranges from southern California (Riverside Co., Fig. 37) to British Columbia.
Fig. 37.

Epargyreus clarus clarus (blue, top), E. c. californicus (red, middle), and E. huachuca (purple, below). Specimens discussed are labeled in color.
Fig. 36.

Epargyreus specimens from the northwestern US (dorsal: left, ventral: right) collected approximately one century ago and their labels: Epargyreus clarus clarus from USA: Oregon (above the line) and Epargyreus clarus californicus from USA: Washington (below the line). All images are to scale, including labels.
Fig. 38.

Epargyreus clarus californicus nectaring on giant vetch in Canada: British Columbia, Cortes Island, 50.0239N 124.9817W, 1-Jun-2019 © Christian Gronau (with permission).
The first record of E. clarus on Cortes Island was a photograph of an adult taken in June 2014 by C. Gronau, and he and Barry Saxifrage have thoroughly documented the use of giant vetch as the larval foodplant (first record of it as a foodplant for E. clarus), through oviposition observations, finding many eggs and larvae, and rearing larvae from eggs. Black locust occurs on Cortes Island, but Gronau and Saxifrage have not found any evidence it is used as a foodplant. Cortes Island is about 320 km north of the Seattle area, Washington where the nearest E. clarus presently occur, and about 520 km from the nearest extant E. clarus clarus populations in southeastern British Columbia. Most Seattle area records are E. clarus clarus, with some known to be temporary introductions from larvae brought in on nursery stock of black locust (Robinia pseudoacacia L.) from eastern North America; however, some records are of E. clarus californicus that may be either migrants from the south or may reproduce locally on an unknown foodplant (Jonathan P. Pelham, pers. comm.). Giant vetch is widespread in the Seattle area; hence, it (or other vetch species) is a potential foodplant for resident populations of E. clarus californicus.
Monoca Grishin, new subgenus
http://zoobank.org/0986D5E1-30DC-46DE-8E66-38F74DA35FD6
Type species. Tagiades monophthalma Plötz, 1884.
Definition. A distant member of the genus Ocella Evans, 1953 (type species Cyclosemia albata Mabille, 1888) (Fig. 39), therefore is defined as a subgenus. Keys to E.26.3 in Evans (1953). Differs from other Ocella species by its single (not double) forewing eyespot (i.e., eyespot only at the end of discal cell and no eyespot in cell CuA1-CuA2), more produced hindwing tornus, straighter (not convex) outer margin of both wings (Fig. 40), and uncus strongly downturned in lateral view.
Fig. 39.

Subgenus Monoca subgen. n. (red) within other Ocella (blue) and more distant relatives.
Fig. 40.

Ocella (Monoca) monophthalma from Brazil: SP, São Bento do Sapucaí. iNaturalist observation 104030146 © rick_costa. CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Etymology. The name is a feminine noun in the nominative singular, given for the single eyespot of the type species: Mono+[O]c[ell]a.
Species included. Only the type species.
Parent taxon. Genus Ocella Evans, 1953.
Comment. Monophyly of this new subgenus with Ocella is weakly supported (Fig. 39, 0.47) and it is possible that it may not hold, in which case Monoca subgen. n. would become a genus-level taxon. Conservatively, due to its monotypy and phenotypic similarities with Ocella, it is proposed as a subgenus in this work.
Pellicia brunneata Williams & Bell, 1939 is a junior subjective synonym of Nisoniades inca (Lindsey, 1925), reinstated status
Sequencing of the holotypes of Pellicia inca Lindsey, 1925 (type locality Peru: Puerto Bermúdez, NVG-22043E01), currently a junior subjective synonym of Nisoniades mimas (Cramer, 1775) (type locality in Suriname) and of Pellicia brunneata Williams & Bell, 1939 (type locality in French Guiana, NVG-15097B04), currently a valid species of Nisoniades Hübner, [1819] (type species Papilio bromius Stoll, 1787, a junior subjective synonym of Papilio mimas Cramer, 1775), reveals that they are most likely conspecific, e.g., their COI barcodes are 100% identical, but differ from their closest relatives, including N. mimas (Fig. 41). Therefore, we reinstate Nisoniades inca (Lindsey, 1925), stat. rest. as a species and place Pellicia brunneata Williams & Bell, 1939, syn. nov. as its junior subjective synonym.
Fig. 41.

Nisoniades inca (red), N. mimas (blue) and relatives.
Abdomen is excluded from the holotype of Staphylus ascalon (Staudinger, 1876)
Genomic sequencing of a leg from the holotype of Helias ascalon Staudinger, 1876 (type locality Brazil: Rio de Janeiro, Nova Friburgo), male, currently in the genus Staphylus Godman and Salvin, 1896 (type species Helias ascalaphus Staudinger, 1876) and a syntype of Staphylus anginus Schaus, 1902 (type locality Brazil: Rio de Janeiro, Nova Friburgo) confirms their synonymy suggested by Mielke (1975). Their COI barcodes are 100% identical and they are collected at the same locality. As Mielke (1975) noted, he dissected an abdomen of a different species (larger size, asymmetrical genitalia) glued to the body of the S. ascalon holotype. To preserve the current usage of the name ascalon and thus to stabilize nomenclature, under ICZN Code Art. 73.1.5, we exclude the abdomen with all its content, including genitalia (dissected by Mielke, in a vial pinned next to the specimen) from the holotype. The COI barcode sequence of the S. ascalon holotype, DNA sample NVG-15033G04, GenBank OP231467, is: AACTTTATATTTTATTTTTGGTATTTGATCAGGAATAGTAGGAACTTCTTTAAGTATTCTTATTCGTTCTGAATTAGGAACTCCTGGATCTTTAATTGGAGATGATCAAATTTATAATACTATTGTAACTGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCTATTATAATTGGAGGATTTGGAAATTGACTTGTACCACTTATATTAGGAGCTCCTGATATAGCTTTCCCTCGTATAAATAATATAAGTTTTTGATTATTACCCCCATCTTTAATACTTTTAATTTCAAGTAGTATTGTAGAAAATGGAGCAGGAACTGGATGAACTGTATATCCCCCACTTTCAGCTAATATTGCCCATCAAGGATCATCAGTAGATTTAGCTATTTTTTCACTTCATTTAGCAGGTATTTCTTCAATTTTAGGAGCAATTAATTTCATTACAACTATTATTAATATACGAATTAATAATTTATCATTTGATCAAATACCTTTATTTGTATGAGCTGTAGGAATTACAGCATTACTTTTACTTTTATCTTTACCAGTATTAGCAGGTGCTATTACTATATTATTAACTGACCGGAATCTTAATACATCATTTTTTGATCCAGCTGGAGGAGGAGATCCTATTTTATATCAACATTTATTT
Xenophanes ruatanensis Godman & Salvin, 1895 is a species distinct from Xenophanes tryxus (Stoll, 1780)
Genomic sequencing of Xenophanes tryxus (Stoll, 1780) (type locality in Suriname) specimens across its range reveal a split into North and South American clades (Fig. 42, tree built from the Z chromosome). The two clades are differentiated genetically with Fst/Gmin of 0.26/0.048 and COI barcode difference between specimens from Costa Rica (NVG-7906) and Guyana (NVG-19088G05) of 1.7% (11 bp) and therefore represent two distinct species. The South American species is X. tryxus, and the oldest name available for the North American populations is Xenophanes ruatanensis Godman & Salvin, 1895, stat. rest. (type locality Honduras: Roatán Island), which we reinstate from synonymy with X. tryxus as a species-level taxon. Genomic sequence of the holotype of Xenophanes perplexus Bell, 1942 (type locality in Mexico: Guerrero) places it as a junior subjective synonym of Xenophanes ruatanensis as expected from its locality, and sequencing of a syntype of Leucochitonea euphemie Ehrmann, 1907 (type locality Venezuela: Suapure) supports its synonymy with X. tryxus (Fig. 42). By default, we consider Hesperia salvianus Fabricius, 1793 (type locality “Indiis”) to be a South American taxon and thus a junior subjective synonym of X. tryxus.
Fig. 42.

Xenophanes ruatanensis (red, above) and Xenophanes tryxus (blue, below). Primary type specimens are labeled in corresponding colors.
Antina Grishin, new genus
http://zoobank.org/5DD0A8EC-EC80-469E-BFDB-1D1056D06623
Type species. Antigonus minor O. Mielke, 1980.
Definition. Genomic sequencing of two paratypes of Antigonus minor O. Mielke, 1980 (type locality in Brazil: Rio Grande do Sul) in ZSMC reveals that they are not monophyletic with Antigonus Hübner, [1819] (type species Urbanus erosus Hübner, [1812]), but instead form a distinct clade that is distant sister to Trina Evans, 1953 (type species Helias geometrina C. Felder & R. Felder, 1867) (Fig. 43). Due to genetic differences and morphological distinction, we do not place A. minor in Trina, but erect a new genus for this unique phylogenetic lineage. In wing pattern and shape (Fig. 44) similar to some species formerly in Antigonus, but currently in Systaspes Weeks, 1901 (type species Antigonus corrosus Mabille, 1878), and subgenus Tiges Grishin, 2022 (type species Antigonus liborius Plötz, 1884) of Paches Godman & Salvin, 1895 (type species Pythonides loxus Westwood, 1852). The similarity is in the two concavities at the outer margin near the hindwing apex, brown wings crossed by two or three dark-brown bands, and small hyaline spots in the middle of postdiscal area of the forewing, valva with a long protuberance from the ampulla, which is particularly bulky and large in this new genus, and not thin and elongated, but thick-C-shaped, curved towards the ventral side of the harpe. Also differs in the paler thin band from near the apex through the middle of the hindwing (Fig. 44). In DNA, a combination of the following base pairs in the nuclear genome is diagnostic: aly2850.4.9:C216T, aly536.218.1:A99T, aly88.17.3:A156G, aly1432.8.2:C101G, and aly4333.3.4:C133T.
Fig. 43.
Antina gen. n. (red), Antigonus (blue), Trina (purple), and Pyrgus (green) among valid genera of Pyrgini.
Fig. 44.

Antina minor from Argentina: Misiones, Candelaria. iNaturalist observations 64610972 (left, color-corrected) and 64761384 (right, rotated) © Patricio A. Mantinian. CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Etymology. The name is a feminine noun in the nominative singular, a fusion of Ant[igonus] + [Tr]ina. The type species is common in A[rge]ntina, where the photographs of live individuals were taken (Fig. 44).
Species included. Only the type species.
Parent taxon. Tribe Pyrgini Burmeister, 1878.
Nervia niveostriga (Trimen, 1864), new combination
Despite the lack of wing pattern typical for Nervia Grishin, 2019 (type species Hesperia nerva Fabricius, 1793), brown and rather humbly patterned Pamphila? niveostriga Trimen, 1864 (type locality in South Africa) is not monophyletic with Kedestes Watson, 1893 (type species Hesperia lepenula Wallengren, 1857) and instead originates within Nervia being sister to phenotypically similar Nervia wallengrenii (Trimen, 1883) (type locality in South Africa) (Fig. 45). Therefore, we propose Nervia niveostriga (Trimen, 1864) comb. nov.
Fig. 45.
Nervia (red) including Nervia niveostriga, and Kedestes (blue).
Leona lota Evans, 1937, reinstated combination and Leona pruna (Evans, 1937) with Leona reali (Berger, 1962), new combinations
Genomic sequencing of Lennia lota (Evans, 1937) (type locality in Cameroon) reveals that it is not monophyletic with the genus Lennia Grishin, 2022 (type species Leona lena Evans, 1937) where it was placed without DNA sequence data (Zhang et al. 2022b), but instead belongs to Leona Evans, 1937 (type species Hesperia leonora Plötz, 1879) as originally proposed, implying Leona lota Evans, 1937, comb. rest. (Fig. 46). Furthermore, sequencing of the holotype of Caenides na Lindsey & Miller, 1965 (type locality in Liberia), currently treated as a junior subjective synonym of Pteroteinon reali Berger, 1962 (type locality in Ivory Coast), reveals that it belongs to the genus Leona (Fig. 46). Therefore, and due to phenotypic similarities with Pteroteinon pruna Evans, 1937 (type locality Cameroon), we propose Leona pruna (Evans, 1937), comb. nov. and Leona reali (Berger, 1962), comb. nov.
Fig. 46.

Leona (red, above) with Leona reali (represented by the holotype of Caenides na) and Leona lota, and Lennia (blue, below).
Lotongus calathus (Hewitson, 1876) complex consists of five species-level taxa
Out of a total of 6 (including the nominal), 5 valid subspecies of Eudamus calathus Hewitson, 1876 (type locality in Sumatra), the type species of and currently in the genus Lotongus Distant, 1886, are differentiated from each other genetically at the level typical for distinct species (Fig. 47). Phenotypic differences between them have been given in detail (Evans 1949) (Fig. 48). Therefore, we propose that they are species-level taxa distinct from the nominotypical L. calathus: Lotongus shigeoi Treadaway & Nuyda, 1994, stat. nov. (type locality in Philippines), Lotongus balta Evans, 1949, stat. nov. (type locality Myanmar: Kanbauk), Lotongus zalates (Mabille, 1893), stat rest. (type locality in Java), and Lotongus taprobanus (Plötz, 1885), stat rest. (type locality in Sulawesi). However, Hesperia parthenope Plötz, 1886 (type locality in Nias), is more closely related to the nominotypical L. calathus (Fig. 47): e.g., COI barcode difference between them is 0.8% (5 bp) compared to 3.3% (22 bp) and 3.8% (25 bp) difference between the nominotypical and the two closest to it species L. balta and L. zalates, respectively (Fig. 47 mitogenome tree). Therefore, we keep Lotongus calathus parthenope (Plötz, 1886), confirmed status, as a subspecies pending further investigations.
Fig. 47.

Lotongus species: balta (green), calathus (red, its subspecies parthenope labeled in magenta), zalates (purple), shigeoi (orange), and taprobanus (blue). Z chromosome (left) and mitogenome (right) trees are shown, leaf order is the same.
Fig. 48.

Lotongus species: a, b. balta, c. calathus, and d. taprobanus. iNaturalist observations: a. 24122671 Thailand: Phetchabun, Khao; b. 23269969 Thailand: Ranong, Kra Buri; c. 63226055 Indonesia, Bangka Isl.; and d. 39632233 Indonesia: Sulawesi, Bolaang Mongondow. c. © Yunita Lestari, others © Les Day. Several images are color-corrected, rotated, and/or flipped. CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Carystus tetragraphus Mabille, 1891 is confirmed as a junior subjective synonym of Lotongus calathus parthenope (Plötz, 1886)
Sequencing its syntype, we tentatively placed Carystus tetragraphus Mabille, 1891 (type locality “Amboine”) as a junior subjective synonym of Lotongus calathus parthenope (Plötz, 1886) (type locality Nias) without genomic data about the latter taxon (Zhang et al. 2022b). Here, we report sequencing of two Hesperia parthenope syntypes confirming this hypothesis (Fig. 47). In the genomic tree, the syntypes of the two taxa group closely together and with a female labeled from “Dutch New Guinea” (i.e., Indonesia: Western New Guinea), and their COI barcodes are 100% identical. The two L. parthenope syntypes, both females, we found in MFNB and sequenced are currently not labeled as types. However, they are from the Weymer collection, labeled in Weymer’s handwriting, collected on Nias in 1883 (prior to the original description in 1886), and one of them (NVG-21118H09), which matches the original description in every detail, bears a label [ parthenope Wm. i l | Plötz StettZeit 1886. ], the last line referring to the published work (Plötz 1886). Therefore, we determine that these specimens are syntypes of L. parthenope.
Parnara bipunctata Elwes & J. Edwards, 1897 is a junior subjective synonym of Borbo impar ceramica (Plötz, 1886), new status and new combination
Genomic analysis (Fig. 49) of a syntype of Hesperia ceramica Plötz, 1886 (type locality in Indonesia: Seram Island) (Fig. 50a) that was illustrated by Ribbe (1889) in his Fig. 6 on Taf. V, reveals that it is not monophyletic with Pelopidas agna larika (Pagenstecher, 1884) (type locality in Indonesia: Ambon Island) where it was placed by Evans (1949) as a junior subjective synonym, but instead is sister to the holotype of Parnara bipunctata Elwes & J. Edwards, 1897 (type locality in Indonesia: Bacan Island) (Fig. 50b), currently a subspecies of Borbo impar (Mabille, 1883) (type locality in Australia or Oceania) (Zhang et al. 2022b). Being phenotypically similar, originating from nearby localities, and having COI barcodes only 0.5% (3 bp) different, the two type specimens likely represent the same taxon that belongs to the genus Borbo Evans, 1949 (type species Hesperia borbonica Boisduval, 1833). Therefore, by the priority of names, we treat Borbo impar ceramica (Plötz, 1886), stat. nov., comb. nov. as a valid subspecies, and propose that Parnara bipunctata Elwes & J. Edwards, 1897 is its junior subjective synonym.
Fig. 49.

Borbo impar ceramica (magenta and red, above) and Pelopidas agna (blue, below) among their relatives.
Fig. 50.

Borbo impar ceramica: a. lectotype ♀ of Hesperia ceramica (Taf. V Fig. 6 from Ribbe (1889) with this specimen reproduced on the right, reduced) and b. holotype ♂ of Parnara bipunctata (Pl. XIX fig. 5 from Elwes & Edwards (1897) with this specimen reproduced on the right, reduced) and their labels (larger scale bar for specimens and smaller one for labels).
Plötz (1886) referred to Ribbe in his original description of H. ceramica, and Ribbe (1889) later wrote that he had collected only one specimen. Hence, this specimen (Fig. 50a) is the best name bearer of the taxon and may be the only syntype. It is not the holotype, because the original description did not state or imply that only one specimen was involved (ICZN Code Art. 73.1.2. and Recommendation 73F). To enhance the stability of nomenclature, N.V.G. hereby designates the sequenced syntype NVG-22016H12 (Fig. 50a) in the Zoologische Staatssammlung München, Germany, bearing the following five rectangular labels, the third purple and others white, the second label agrees with Plötz’s handwriting: [ Ceram | Jllo | C.Ribbe 1884 ], [Hesperia | Ceramica Pl. ], [ Original ], [ ♀ Parn. ceramica Pl. | typ. (sec Mab=plebeia) | Ceram ], and [ DNA sample ID: | NVG-22016H12 | c/o Nick V. Grishin ] as the lectotype of Hesperia ceramica Plötz, 1886. The specimen of P. bipunctata we sequenced (NVG-18074H04 in MFNB, Fig. 50b) is the holotype by monotypy, because the original description explicitly stated that this species was described from one specimen (Elwes and Edwards 1897).
Oxynthes martius (Mabille, 1889), reinstated status and South America as a likely type locality of Oxynthes corusca (Herrich-Schäffer, 1869)
Genomic sequencing of a syntype of Goniloba corusca Herrich-Schäffer, 1869 (type locality not specified), the type species of its current genus Oxynthes Godman, 1900, reveals that the syntype groups with specimens from South America, including the holotype of Xeniades leucogaster Röber, 1925 (type locality in Brazil: Rio Grande do Sul), currently considered a junior subjective synonym of O. corusca following Evans (1955) (Fig. 51). These South American specimens are well-separated in the tree from North American specimens identified as O. corusca, including the holotype of Proteides martius Mabille, 1889 (type locality Panama: Chiriqui) that is currently treated as a junior subjective synonym of O. corusca. The COI barcode difference between the syntype of O. corusca and the holotype of P. martius Mabille, 1889 is 3.6% (24 bp) suggesting that they are distinct species. Furthermore, Fst/Gmin statistics between the two clades are 0.36/0.04. First, to ensure nomenclatural stability and unambiguous identification of O. corusca, N.V.G. hereby designates its sole syntype in MFNB bearing the following eight rectangular labels, the first one lilac-colored and others white: [ Origin. ], [ corusca HS. ], [ Coll. H.–Sch. ]. [ Coll. Staudgr. | Kasten 671], [ Coll. | Staudinger ], [ Corusca | H-Sch. ], [ {QR code} http://coll.mfn-berlin.de/u/ | 3226be ], and [ DNA sample ID: | NVG-15035C03 | c/o Nick V. Grishin ] as the lectotype of Goniloba corusca Herrich-Schäffer, 1869. The lectotype is lacking the abdomen and both antennae. Second, because the lectotype is in the clade consisting of South American specimens (Figs. 51 blue, 52 right) that is separate from the clade of North American specimens (Figs. 51 red, 52 left), we suggest that the type locality of O. corusca is in South America. Third, due to genetic differentiation, we propose species-level status for Oxynthes martius (Mabille, 1889), stat. rest.
Fig. 51.

Oxynthes martius (red, above) and O. corusca (blue, below). Primary types are labeled in corresponding colors.
Fig. 52.

Oxynthes martius (left) and O. corusca (right). iNaturalist observations 124556487 Mexico: Oaxaca © John Kemner and 34915995 Brazil: Mato Grosso, Alta Floresta, © belgianbirding, respectively, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Notamblyscirtes durango J. Scott, 2017, new status
Phylogenetic analysis of Notamblyscirtes Scott, 2006 (type and the only species Amblyscirtes simius W. H. Edwards, 1881) from across the range reveals their partitioning into two well-separated clades (Fig. 53). The first clade (Fig. 53 blue) includes the lectotype of N. simius (type locality in USA: Colorado, Pueblo Co.). The second clade (Fig. 53 red) contains the holotype of Notamblyscirtes simius durango J. Scott, 2017 (type locality in Mexico: Durango), proposed as a subspecies. Specimens from southeastern Arizona are in this red clade, however, those from eastern Arizona (the White Mountains) are in the blue clade. The two clades likely represent two distinct species, rather than subspecies, due to their genetic differentiation reflected in Fst/Gmin statistics of 0.49/0.009 and in 2% (13 bp) COI barcode difference between the primary type specimens of the two taxa. Differences in their phenotypes, such as the darker appearance and a different pattern of forewing apical spot in N. simius durango, agree with this genetic differentiation. Therefore, we propose species-level status for Notamblyscirtes durango J. Scott, 2017, stat. nov.
Fig. 53.

Notamblyscirtes durango (red, above) and N. simius (blue, below). Primary types are labeled in corresponding colors.
Rhinthon? zaba Strand, 1921 is a junior subjective synonym of Conga chydaea (A. Butler, 1877), not of Cynea cynea (Hewitson, 1876)
Sequencing of the holotype of Rhinthon? zaba Strand, 1921 (type locality Mexico: Veracruz, Orizaba, NVG-20082H04), currently a junior subjective synonym of Cynea cynea (Hewitson, 1876) (type locality in Venezuela, subtribe Moncina A. Warren, 2008), reveals that the two taxa are in different subtribes, and the former is placed within specimens of Conga chydaea (A. Butler, 1877) (type locality Brazil: Amazonas, Serpa, subtribe Hesperiina Latreille, 1809) (Fig. 54). COI barcodes of R. zaba and C. chydaea (NVG-18119D10) are 100% identical and the specimens are phenotypically similar. Therefore, we propose that Rhinthon zaba Strand, 1921 is a junior subjective synonym of Conga chydaea (A. Butler, 1877), a new synonym placement. This situation is the same as with Pamphila binaria Mabille, 1891 (type locality Venezuela) that we identified previously (Zhang et al. 2022b) as a junior subjective synonym of C. chydaea and not of C. cynea.
Fig. 54.

Conga chydaea (red with Rhinthon zaba labeled in magenta) and Cynea cynea (blue) among their relatives. Primary types are labeled in corresponding colors.
Genomic revision of Hedone Scudder, 1872
A phylogenetic tree constructed from protein-coding regions in the Z chromosome of Hedone Scudder, 1872 (type species Hesperia brettus Boisduval & Le Conte, [1837], which is a junior subjective synonym of Hedone vibex vibex (Geyer, 1832)) specimens across the Americas suggests that the genus consists of eight species, judging by the number of strongly supported prominent clades. Each such clade of more than one specimen has an appearance of a comb rather than a well-resolved tree (Fig. 55). Such clade shape is consistent with it representing a species: a set of populations that exchange genes across the range. Out of these eight clades, three have been treated as species distinct from H. vibex (type locality “West Indies”, likely in error), namely, Hedone dictynna (Godman & Salvin, 1896) (type locality St. Vincent & Grenada), Hedone bittiae (Lindsey, 1925) (type locality Peru: Matucana), and Hedone vibicoides (de Jong, 1983) (type locality Suriname: Zanderij). Out of the remaining four clades, one corresponds to a new species described below, and three correspond to taxa currently considered subspecies of H. vibex. Out of these, Hesperia catilina Plötz, 1886 (type locality Brazil: Santa Catarina, Blumenau; syntype NVG-18052B01 sequenced) is the most distinct genetically and may not be monophyletic with H. vibex. Support for the species-level distinction between H. vibex and Hedone praeceps Scudder, 1872 (type locality Mexico: Oaxaca, Tehuantepec) (Fig. 56) is given in Cong et al. (2019a): Fst/Gmin statistics are 0.84/0.001 (Cong et al. 2019a) and COI barcodes differ by 3.2% (21 bp). Finally, Polites vibex calla Evans, 1955 (type locality Peru: Callao) is sister to H. praeceps and differs from it by 2.7% (18 bp) in the COI barcode. Therefore, we reinstate two taxa as species: Hedone praeceps Scudder, 1872, stat. rest. and Hedone catilina (Plötz, 1886), stat. rest., and propose to treat Hedone calla (Evans, 1955), stat. nov. as a species-level taxon. We note that Hesperia catilina Plötz, 1886 is a junior primary homonym of Hesperia catilina Fabricius, 1793, currently regarded as a valid subspecies of Leptotes cassius (Cramer, 1775). However, because both names have not been considered congeneric after 1899, under Art. 23.9.5 of the ICZN Code (1999), the case should be referred to the ICZN for a ruling under the plenary power. Therefore, in the interest of the stability of nomenclature, we use Hedone catilina instead of proposing a new name.
Fig. 55.
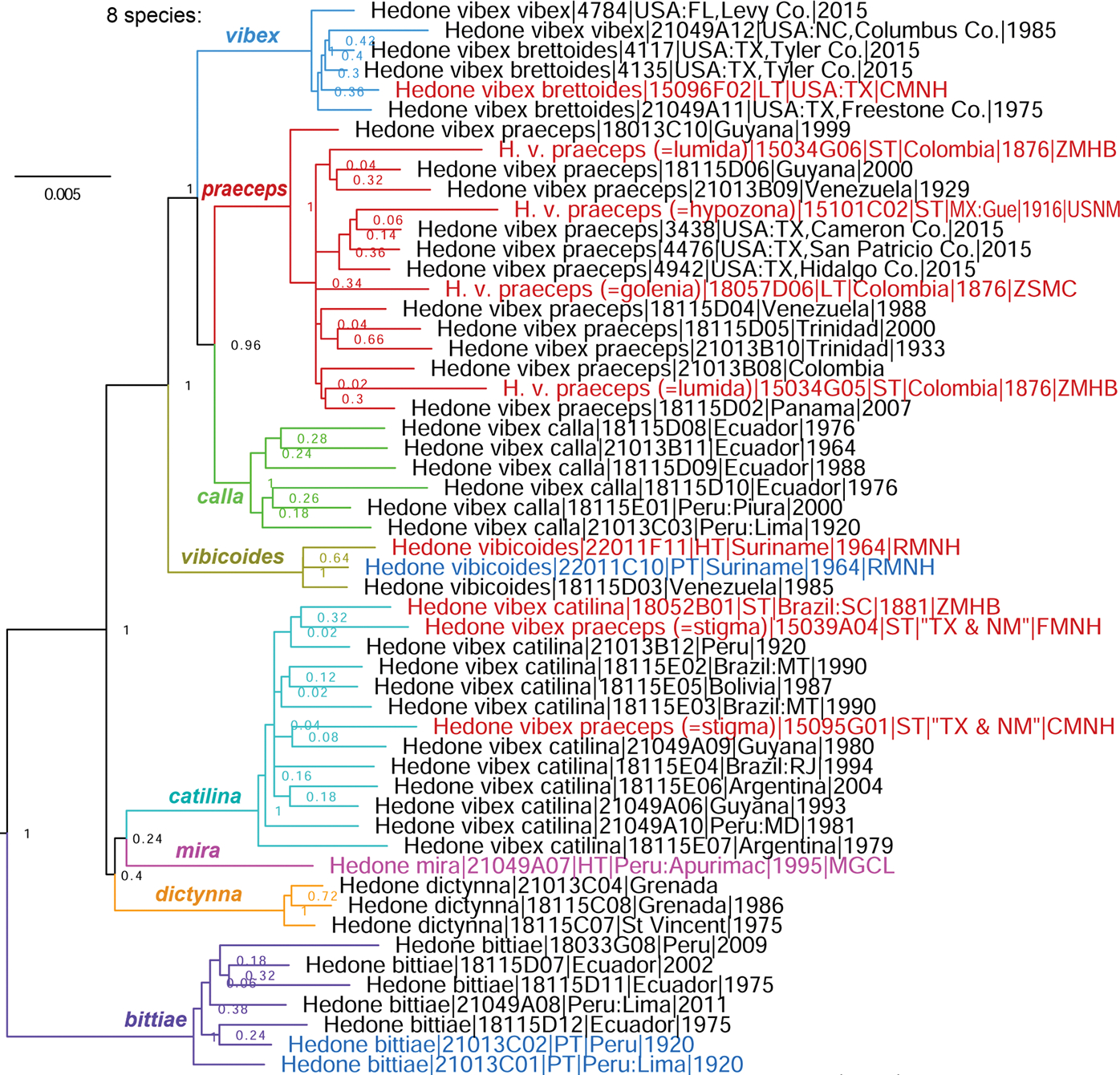
Clades for 8 species of Hedone are shown in different colors and labeled with species names. Primary and secondary types are labeled in red and blue respectively, except that the holotype of Hedone mira sp. n. is in magenta.
Fig. 56.

Females of the two USA Hedone species: a, b. vibex, and c, d. praeceps. iNaturalist observations: a. 133774978 GA, Liberty Co. © Tom Austin; b. 109723959 FL, Charlotte Co. © Jay Horn; and TX: Hidalgo Co.: c. 109408293 McAllen © ronthill and d. 103375027 Mission © James Bailey. Several images are color-corrected, rotated, and/or flipped. CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
The genomic tree reveals a curious genetic homogeneity of specimens within each species, even over the vast ranges (Fig. 55). E.g., specimens of H. praeceps from Texas (Fig. 56c, d) and Guyana reveal limited genetic differentiation, while Guyanese specimens of H. praeceps and H. catilina differ very significantly from each other. Finally, as a surprise, we find that two syntypes of Pamphila stigma Skinner, 1896 (type locality: USA: southern New Mexico and southwest Texas), one from FMNH and the other from CMNH, are not monophyletic with H. praeceps, a synonym of which P. stigma currently is, but belong to H. catilina, a South American taxon. Wing patterns agree with this placement. Provided that P. stigma syntypes in two different collections are H. catilina, we place P. stigma as a junior subjective synonym of H. catilina and deduce that these syntypes were mislabeled. The type locality of P. stigma becomes “South America” and figuring it out more precisely by genomics will require additional sequencing and analysis. This example illustrates that care needs to be taken when assigning synonymies by reported type localities and not by the type specimens themselves, even if reported localities seem believable. However, in this case, the locality does cause suspicions, because currently no Hedone species is known from around “southern border of New Mexico and S.-W. Texas”.
Hedone mira Grishin and Lamas, new species
http://zoobank.org/365340C5-AF20-4B5F-975C-6F6975C25C0A
Fig. 57.

Holotype of Hedone mira sp. n. dorsal (left) and ventral (right) views, NVG-21049A07, data in text.
Definition and diagnosis. enomic sequencing of Hedone Scudder, 1872 (type species Hesperia brettus Boisduval & Le Conte, [1837], which is a junior subjective synonym of Hedone vibex vibex (Geyer, 1832)) specimens revealed one that didn’t group with any known taxa (Fig. 55 magenta) and therefore was probably a new species. Inspection of the phenotype of this specimen (Fig. 57) did not suggest a possible match to species currently placed in other genera. This new species confidently belongs to Hedone and has a unique appearance making its identification straightforward. It differs from all other Hesperiidae species by a combination of the following characters: antenna about half of costa length, with thick club; stigma broad, typical of Hedone, encircled by extensive patches of dark scales; ventral forewing mostly orange-yellow, nearly not darkened in the middle, with brown band along outer margin, two brown elongated spots near the apex, brown base and inner margin; ventral hindwing mostly rusty-brown with broad orange-yellow ray along 1A+2A vein, cream-colored spot at the end of discal cell, cream-orange spot in the middle of cell Sc+R1-RS, and a cream-orange postdiscal band of five spots (separated by darker veins) between veins M1 and 1A+2A. In summary, it looks very much like Hedone bittiae (Lindsey, 1925) (type locality in Peru) on dorsal side, but ventrally the wing pattern is quite different: the hindwing is mostly brown with orange spots rather than yellow with brown spots.
Barcode sequence of the holotype: Sample NVG-21049A07, GenBank OP231472, 658 base pairs: AACTTTATATTTTATTTTTGGTATTTGAGCAGGAATATTAGGAACTTCTTTAAGTCTATTAATTCGAACAGAATTAGGTAATCCTGGCTCTTTAATTGGAGATGATCAAATTTATAATACTATTGTAACGGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCTATTATAATTGGAGGATTTGGAAATTGATTAGTTCCTTTAATATTAGGAGCTCCTGATATAGCTTTTCCTCGAATAAATAATATAAGATTTTGAATATTACCCCCTTCACTAACATTATTAATTTCAAGAAGAATTGTAGAAAATGGTGCAGGAACAGGTTGAACAGTTTATCCACCTTTATCTTCTAATATTGCTCATCAAGGATCTTCTGTTGATTTAGCAATTTTTTCTCTTCATTTAGCTGGAATTTCTTCTATTTTAGGAGCCATTAACTTTATTACAACAATTATTAATATACGAATTAAAAATTTATCTTTTGATCAAATACCTTTATTCGTATGATCTGTTGGAATTACAGCTCTATTATTATTATTATCTTTACCTGTTTTAGCTGGAGCTATTACTATATTACTTACAGATCGAAATTTAAATACTTCTTTTTTTGATCCAGCTGGAGGAGGAGATCCAATTTTATATCAACATTTATTT
Type material. Holotype: ♂ deposited in the McGuire Center for Lepidoptera and Biodiversity, Gainesville, FL, USA (MGCL), bears four rectangular printed labels: three white [ SÜDPERU, Apurimac | Jarjatera, 30 km von | Abancay, 2800m | 5.XI.-22.XII.1995 | RAINER MARX leg. | EMEM, 8.X.1995 ], [ W. McGuire colln. | MGCL Accession | # 2008–43 ], [ DNA sample ID: | NVG-21049A07 | c/o Nick V. Grishin ], and one red [ HOLOTYPE ♂ | Hedone mira | Grishin and Lamas ]. Paratypes: 9 ♂♂, 2♀♀, all from Peru in MUSM, [Huancavelica department]: 1 ♂, road [Nueva Esperanza de] Chonta [12° 37’ S, 74° 29’ W] - Churcampa [12° 44’ S, 74° 23’ W], 3000 m, 3-Dec-2000, leg. V. Doroshkin; Apurímac department: 1 ♂, 7 km NWW Chalhuanca, 14° 13’ S, 73° 19’ W, 2770 m, 16-Mar-1987, leg. O. Karsholt; 1 ♀, Quebrada Pacpapata, 13° 40’ S, 72° 55’ W, 1830 m, 2-Oct-2004, leg. J. Grados; Cuzco department: 2 ♂♂, Calca, [13° 19’ S, 71° 57’ W], 2950 m, 5-Aug-1983, leg. J. L. Venero; 1 ♂, same data, but 8-Aug-1983; 2 ♂♂, cerca [= near] Pisac [13° 25’ S, 71° 51’ W, 3100m], 22-Sep-1989, leg. N. Jara; 1 ♀, Chocco [13° 33’ S, 71° 59’ W, 3450m], 25-Nov-1988, leg. G. Valencia; 1 ♂, Quebrada Uraca, SW of Limatambo, 13° 30’ S, 72° 28’ W, 2800 m, 21-Jun-2003, leg. G. Lamas; 1 ♂, same data, but leg. C. Peña.
Type locality. Peru: Apurímac department, Carcatera area, 30 (road) km NW from Abancay, el. 2800 m.
Etymology. The name is given for the unexpected for the genus appearance of the ventral wing pattern of this species: mira[culous], a Latin word for astonishing, extraordinary, wonderful. The name is a feminine noun in the nominative singular.
Distribution. Recorded from Huancavelica, Apurímac, and Cuzco departments in southern Peru.
Hesperia peckius W. Kirby, 1837 is a junior subjective synonym of Polites (Polites) coras (Cramer, 1775)
Here, we argue that information we assembled about Papilio coras Cramer, 1775 (type locality Suriname) is sufficient to unambiguously associate this name with a known species. The illustrations and the description of P. coras refer to female(s) of a North American species currently known as Polites (Polites) peckius (W. Kirby, 1837), and we arrived at this conclusion with the following arguments.
Cramer volumes (1775–1780), where the original description of P. coras was published, contain Dutch text in the left column and French text in the right column. The two texts are similar and differ only in small details. The Dutch description of P. coras is: “Dit Dikkopje (Papilio Urbicola) is wederzyds van dezelfde bruine kleur, met geelagtige vlakken. De sprieten der Kapellen, welken men gewoon is Dikkoppen te noemen, eindigen aan den knop met een haakagtig puntje. Deze en de twee voorgaande berusten in de verzameling van den Heer E. de Marre. Het is uit Surinamen.” We translate it literally as “This small thickhead [=skipper] (Papilio Urbicola) is on both sides of the same brown color, with yellowish patches. The antennae of the butterflies, which one is accustomed to call thickheads, end in the knob with a hook-like point. This and the previous two [species] are in the collection of Mr. E. de Marre. It is from Suriname.” The French text and its literal translation: “Le dessous de ce Plebejen noble ou têtu (Pap. Pleb. Urbicolae) est de la même couleur brune, à taches jaunâtres, que le dessus. Les antennes des Papillons qu’on a coutume de nommer têtus, finissent à la masse en pointe crochue. Celui-ci & les deux precedents se trouvent dans la Collection de Mr. E. de Marre. Il est de Suriname.” “The underside of this commoner [=Plebeian] urban [=noble] [i.e., Plebejus Urbicola] or stronghead [=skipper] (Pap. Pleb. Urbicolae) is the same brown color, with yellowish spots, as the upper surface. The antennae of the butterflies, which we usually call strongheads [=skippers], end in a hooked point on the club. This one & the two preceding ones are in the collection of Mr. E. de Marre. It is from Suriname.”
Importantly, both descriptions state that the ventral side of wings is brown color (as dorsal) and with yellow patches/spots. This information complements the illustrations of the dorsal side, both the published engraving (Fig. 58b right) and the original drawing by G. W. Lambertz (Fig. 58b left). The most unusual (for Hesperiidae) feature of the dorsal illustration is the doublet of long, perfectly aligned dash-like spots in hindwing cells M1-M2 and M2-M3. These spots are longer than the neighboring spots of the postdiscal row and protrude farther from them towards the outer margin than towards the base. First, we looked for Hesperiinae species that would be expected from Suriname and match both the illustrations and the description of P. coras. We critically inspected every known Hesperiinae species for P. coras characters. Some Hylephila Billberg, 1820 (type species Papilio phyleus Drury, 1773) possess the doublet of hindwing spots, but they also have an orange-yellow ray along hindwing vein 1A+2A, which is absent in P. coras, and their ventral side is yellow-orange with brown spots rather than brown with yellow spots. Cantha zara (E. Bell, 1941) (type locality in Bolivia) has the doublet of spots and its wings ventrally are orange-brown with yellow spots, but the two spots in the hindwing doublet are not aligned with each other, and the spot in M2-M3 is offset basad (Fig. 58f). Finally, some females of Vernia dares (Plötz, 1883) (type locality not specified) match P. coras illustration very well, except that their forewing spots are rounder and smaller (Fig. 58g), but they are ventrally yellow with brown spots, not brown with yellow spots. Consistently with our studies, Olaf H. H. Mielke (2005: 1209), the foremost expert in American Hesperiidae, is of an opinion that P. coras does not match any neotropical species.
Fig. 58.

Specimens and original illustrations: a–e, h, i. Polites coras: a. P. c. coras neotype, dorsal (left) and ventral (right) views; b. original drawing by Lambertz (left) and published engraving (right); c–e, h. variation in specimens that are similar to the original illustrations; i. P. coras peckius original illustration, dorsal (left) and ventral (right) views. f. Cantha zara holotype. g. Vernia dares from Mexico: Oaxaca. All specimens are females, except f. Photographs a are by Riley J. Gott, b (left), c–f and h are © The Trustees of the Natural History Museum London and are made available under Creative Commons License 4.0 (https://creativecommons.org/licenses/by/4.0/), and f is by Bernard Hermier.
Therefore, we suspected that the locality “Suriname” was erroneous. This is not without a precedent: Rothschild and Jordan (1903) wrote “it is quite clear that … the localities in E. de Marre’s collection were not reliable.” For example, for the two species described by Cramer previous to P. coras, also from the de Marre’s collection, the “Suriname” locality is apparently incorrect. One of these species is currently regarded as Indomalayan Jamides celeno (Cramer, 1775), and the other is a nomen dubium, Papilio arius Cramer, 1775 (could be either Lycaenidae or Riodinidae), which does not match any known species. Thus, we analyzed all Old World Hesperiinae, in particular from the tribe Taractrocerini Voss, 1952, many of which are small brown, orange-spotted species in the genera Taractrocera Butler, 1870 (type species Hesperia maevius Fabricius, 1793), Potanthus Scudder, 1872 (type species Hesperia omaha W. H. Edwards, 1863), and Telicota Moore, [1881] (type species Papilio colon Fabricius, 1775), among others. Among them, we were not able to find a match to P. coras.
Out of the entire known worldwide Hesperiinae fauna, only some specimens identified as Polites (Polites) peckius (W. Kirby, 1837) (type locality unclear, probably northeastern North America) fit what is known about P. coras except its locality (Fig. 58a, c–e, h, i). To test our suspicion about the P. coras identity, we searched for its primary type specimens. We inspected collections that house possible Cramer types: BMNH and RMNH, and failed to find candidate specimens. Assuming that they are lost, we proceeded with the neotype designation. We believe that there is an exceptional need to designate the neotype, because the identity of this taxon has been questionable for years since its description, essentially resulting in a dual nomenclature, and its type locality is erroneous. Hereby, N.V.G. designates the specimen, a female, shown in Fig. 58a as the neotype of Papilio coras Cramer, 1775.
Our neotype of P. coras satisfies all requirements set forth by the ICZN Article 75.3, namely: 75.3.1. It is designated to clarify the taxonomic identity of Papilio coras Cramer, 1775, which has been questioned since its original description, and to define its type locality; 75.3.2. The characters for the taxon have been quoted from the original description above and include the atypical (for Hesperiinae) doublet of long orange dashes in the middle of the dorsal hindwing and the brown, yellow-spotted ventral wing surface; 75.3.3. The neotype bears three rectangular labels: [ exeggex ♀ | August 10 1987 | Flourtown | Montgomery Co. Pa. | Coll. RW Boscoe // “grasses” | emerged | November 4 1987 ], [ FSCA | Florida State Collection | of Arthropods ], and [ DNA sample ID: | NVG-22051H08 | c/o Nick V. Grishin ]; 75.3.4. Our search for syntypes is described above, was unsuccessful, therefore we believe that they were lost; 75.3.5. The neotype is consistent with the original drawing, published engraving, and the original description. It only differs from the illustrations by the presence of a yellow-orange spot in the hindwing discal cell. However, this spot is variable, and is missing in some specimens (Fig. 58c, d); 75.3.6. The neotype is from USA: Pennsylvania, Montgomery Co., Flourtown, which becomes the type locality of P. coras. The type locality was given as Suriname in the original description, likely by mistake, and the original types could have been collected in the general New York City area, as for some other species described by Cramer, e.g., Heraclides cresphontes (Cramer, 1777); 75.3.7. The neotype is in the collection of the McGuire Center for Lepidoptera and Biodiversity, Gainesville, FL, USA (MGCL).
As a result of this neotype designation, P. coras belongs to the genus Polites Scudder, 1872 (type species Hesperia peckius W. Kirby, 1837) and is likely conspecific with P. peckius. We note that the original illustration of Polites peckius syntype(s) reproduced here as Fig. 58i shows differences in wing patterns from both the illustration and the neotype of Polites coras. In P. peckius, the orange spots are larger, and the hindwing pattern differs. Instead of separate narrow spots, there is a yellow-orange blotch narrowly cut through by dark veins. The two long dashes in cells M1-M2 and M2-M3 are wider and do not sharply stand out from other spots, especially at the basal side of the discal band, which is more even than in P. coras. Although particularly widespread in the upstate New York area (Fig. 59), this phenotype occurs in all northeastern populations and may not be a reflection of geographic variation. Therefore, we propose that Hesperia peckius W. Kirby, 1837, syn. rev. is a junior subjective synonym of Polites (Polites) coras (Cramer, 1775). We keep the subspecies Polites coras surllano J. Scott, 2006 comb. nov. as valid but use the name in a new species-subspecies combination. More detailed analysis of specimens across the range of P. coras, including their phenotypes and genetics, will shed light on its variation, population structure, and rigorous definition of subspecies.
Fig. 59.

Polites coras (peckius-like) from USA: upstate New York. iNaturalist observations: a. 54682030, Saratoga Co. © Steven Auletta; b. 13028519, NY: Monroe Co. © Chris Alice Kratzer; c. 56710259. NY: Tompkins Co. © Brad Walker, edited to fill out the frame and color-corrected from the original. CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
The priority of P. coras over P. peckius may not be welcomed by some lepidopterists, but we think that the evidence presented here is sufficiently strong to support it and to unambiguously identify the Lambertz’s illustration augmented with the ventral side description by Cramer. If the resulting reinstatement of the name coras used prior to 1981 (Draudt 1921–1924; Evans 1955), and used as valid in the recent comprehensive catalogue of the Neotropical Hesperiidae (Mielke 2004; Mielke 2005), is not desirable, ICZN could be requested to intervene.
Pompe Grishin and Lamas, new genus
http://zoobank.org/A5199A2E-BBAF-41B3-AD96-C87B5018AA84
Type species. Lerema postpuncta Draudt, 1923.
Definition. In the same clade with Serdis Mabille, 1904 (type species Serdis flagrans Mabille, 1904, a subspecies of Pamphila statius Ménétriés, 1857) and Cyclosma Draudt, 1923 (type species Cyclosma abdonides Draudt, 1923), but far removed from Pompeius Evans, 1955 (type species Hesperia pompeius Latreille, [1824]) where it was placed previously (Fig. 60). Keys to M.15.3. in Evans (1955). Distinguished from its relatives by a combination of the following characters: stigma narrow, flanked with gleaming scales (Fig. 61); uncus divided, arms diverging, not widely separated; gnathos arms converging, as long as uncus; harpe of valva dorsally expanded into a lobe, distal margin concave, ventrally with a short spike directed posterodorsad; aedeagus terminally enlarged. In DNA, a combination of the following base pairs in nuclear genome is diagnostic: aly671.51.3:T45C, aly1838.49.3:A2211T, aly1222.14.14: A5433G, aly1222.14.14:A5412G, aly1838.49.3:T1965G.
Fig. 60.

Pompe gen. n. (red, above) and Pompeius (blue, below).
Fig. 61.

Pompe postpuncta NVG-18117H03, Brazil: MT.
Etymology. The name is a feminine noun in the nominative singular, derived from Pompeius, the former genus name of the type species.
Species included. Only the type species.
Parent taxon. Subtribe Hesperiina Latreille, 1809.
Atalopedes huron (W. H. Edwards, 1863) is a species distinct from Atalopedes campestris (Boisduval, 1852)
A phylogenetic tree constructed from protein-coding genes in the Z chromosome reveals partitioning of Atalopedes campestris (Boisduval, 1852) (type locality in USA: California, Sacramento Co.) into two prominent clades (Fig. 62) that correspond to distinct species according to Fst/Gmin of 0.61/0.02 and COI barcode difference of 1.4% (9 bp). Moreover, genetic differentiation between Californian specimens of nominotypical A. campestris and all others is comparatively much larger than the differentiation between eastern USA specimens and those from Venezuela. The oldest available name for the nonCalifornian clade (Fig. 62 red) is Hesperia huron W. H. Edwards, 1863 (type locality USA: Illinois, Cook Co., Evanston, neotype NVG-15097H11 sequenced). Due to genetic differentiation, we reinstate Atalopedes huron (W. H. Edwards, 1863), stat. rest. as a species-level taxon.
Fig. 62.
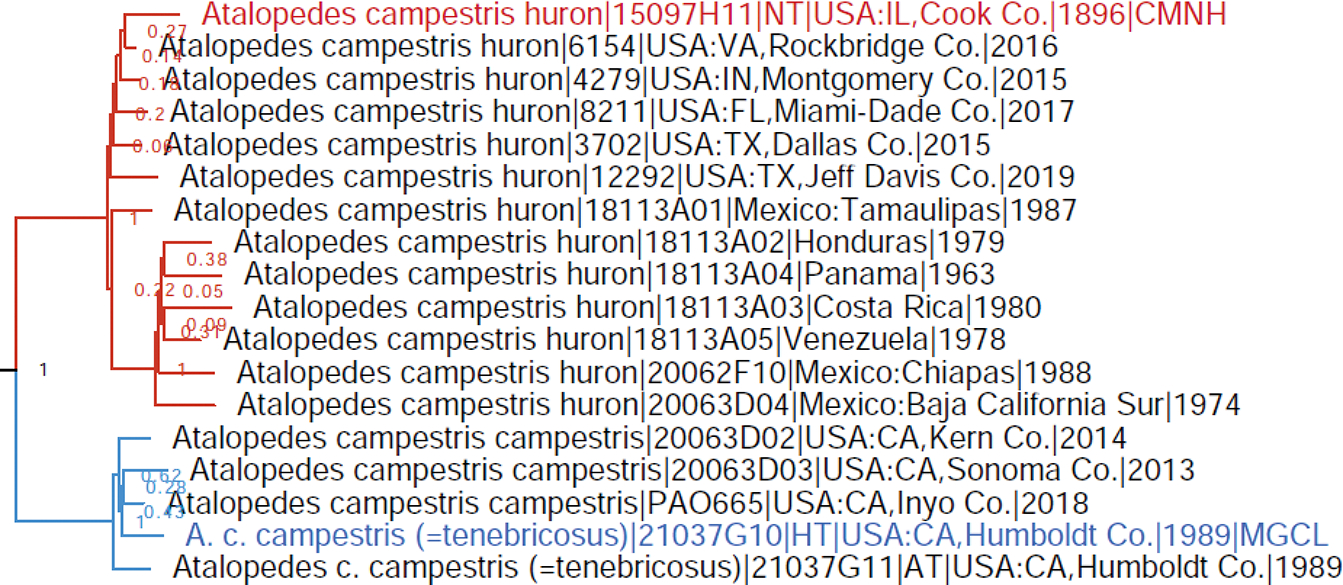
Atalopedes huron (red, above) and Atalopedes campestris (blue, below). Primary type specimens are labeled in corresponding colors.
Putuma Grishin, new subgenus
http://zoobank.org/AB0279C1-5BCE-4A91-A54A-D614A9EF33AF
Type species. Tisias putumayo Constantino and Salazar, 2013.
Definition. Tentatively placed in the subgenus Tixe Grishin, 2022 (type species Cobalus quadrata Herrich-Schäffer, 1869) without genomic data, Tisias putumayo Constantino and Salazar, 2013 is not monophyletic with it and is instead sister to all other Xeniades Godman, 1900 (type species Papilio orchamus Cramer, 1777), including subgenera Cravera de Jong, 1983 (type species Cravera rara de Jong, 1983) and Tixe (Fig. 63). Therefore, it is a subgenus of its own. Distinguished from its relatives by ventral hindwing with a single transverse postdiscal white band from apex to about the middle of cell CuA2-1A+2A, two framed with black bluish spots basad of the band (Fig. 64), and the two hyaline spots in forewing discal cell (in males) being farther from each other than in all other congeners. In DNA, a combination of the following base pairs in nuclear genome is diagnostic: aly420.34.2:C721A, aly1772.2.2:G63A, aly420.34.2:G258A, aly420.34.2:A300C, and aly1931.9.21:G121A.
Fig. 63.

Genus Xeniades with subgenera Xeniades (blue), Cravera (magenta), Tixe (green), and Putuma subgen. n. (red) and genus Oligoria (olive).
Fig. 64.

Xeniades putumayo from Ecuador: Napo, Rio Pingullo. iNaturalist observation 68121597. © Ken Kertell, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Etymology. The name is a feminine noun in the nominative singular, formed from the type species name by removing the last two letters.
Species included. Only the type species.
Parent taxon. Genus Xeniades Godman, 1900.
Lectotype designation for Pamphila trebius Mabille, 1891
To ensure identification of this name, and to stabilize nomenclature, N.V.G. hereby designates a specimen in MFNB shown in Fig. 65 with the following eight rectangular labels, first purple, others white: [ Origin. ], [ Bogota | Nolcken ], [ P. trebius Mb. ], [ Pamph. | Trebius | Mab. ], [ Coll. | Staudinger ], [ Trebius | Mab. ], [ {QR code} http://coll.mfn-berlin.de/u/ | 44a090 ], and [ DNA sample ID: | NVG-15034E04 | c/o Nick V. Grishin ] as the lectotype of Pamphila trebius Mabille, 1891 (type locality Colombia: Bogota). The lectotype is a syntype (possibly the only one in existence, but we are avoiding an assumption of the holotype), because it agrees with the original description (see below), was described in a publication that mentions in the introduction that some of the specimens used were from the Staudinger collection (the syntype is from this collection), one of its identification labels (“P. trebius Mb.”) matches Mabille’s handwriting, and the syntype is curated as a type specimen (“Origin.”).
Fig. 65.

Lectotype of Pamphila trebius Mabille, 1891 dorsal (left) and ventral (right) views, NVG-15034E04, details in text. Wingspan is 27 mm per original description. Larger 1 cm scale bar is for the specimen and smaller one is for labels.
The original description of P. trebius can be (literally) translated as follows (numbers in curly brackets are for the arrows pointing to these characters in Fig. 65): “Pale brown; three small apical dots {1} and another [dot] in the 4th interval {2} white. Hindwings with yellowish hairs. Underside of the forewings brown; veins tinted red {3}, the dots of the upperside a little larger: a small dot additionally in the 3rd interval {4}. Outer margin tinted with lilac gray {5}. Hindwings dusky with anterior portion darker {6}, letting stand out two paler and indistinct square spots in intervals 7 {7} and 8 {8}. Body dusky with red hairs. Palpi dark gray. Antennae with club yellowish below {9}. 27 mm {10} – ♂– Bogota” (Mabille 1891: CLXXIV). We note that Mabille used a different notation of wing cell numbers, presumably i + 1 for i > 1 compared to i in Evans (1953), thus numbering Evans’ forewing cells 1A and 1B as 1 and 2. The apparent wingspan of the lectotype measures 26 mm, however, it is not mounted flat but with wings angled downwards, thus decreasing the actual wingspan. The lectotype designation stabilizes the current treatment of P. trebius as a junior subjective synonym of Cymaenes lumina (Herrich-Schäffer, 1869) (type locality not specified, likely southern parts of South America) (Zhang et al. 2022b). The type locality of P. trebius is questionable because this phenotype is not known from Colombia. As demonstrated by Zhang et al. (2022b), Cymaenes lumina (that includes P. trebius as a synonym) is a South American species, not recorded from North America. A species formerly known as “Cymaenes trebius” that enters the US in South Texas, is Cymaenes isus (Godman, 1900) (type locality Mexico: Guerrero) (Fig. 66). The two species can be distinguished from each other by wing patterns. In C. isus (and Cymaenes edata (Plötz, 1882)), ventral hindwing typically has a pale postdiscal band of spots (separated by darker veins) on brown background, the discal brown area is comparatively narrower, straighter, and the discal spot in cell Sc+R1-RS is usually connected with the basal pale area (Fig. 66 left). In C. lumina, ventral hindwing is overall paler in appearance, with a broader, contrasting discal L-shaped brown patch, and the discal spot in cell Sc+R1-RS is separated from the basal pale area by this brown patch (Figs. 65, 66 right).
Fig. 66.

Cymaenes isus (left) and C. lumina (right). iNaturalist observations 71267802, USA: TX, Hidalgo Co. © Rich Kostecke and 73322525 Argentina: Buenos Aires © Facundo Chieffo, respectively, color-corrected and rotated, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Methionopsis Godman, 1901 is a subgenus of Mnasinous Godman, 1900
Correcting a mistake made in Zhang et al. (2022b), we state that Methionopsis Godman, 1901 is a subgenus of Mnasinous Godman, 1900. These two names were swapped in Zhang et al. (2022b), and this error is corrected here to follow the priority of the two names (1901 vs. 1900). The arguments for their synonymy are the same as presented previously (Zhang et al. 2022b), e.g., COI barcodes of Mnasinous (Methionopsis) modestus (Godman, 1901) and Mnasinous (Mnasinous) patage Godman, 1900 differ by 9.1% (60 bp), while those of Methion melas Godman, 1900 (sister genus, monotypic) differ from them by 12.2–12.3% (80–81 bp).
Mnasinous (Methionopsis) phaeomelas (Hübner, [1829]), new combination
Genomic sequencing of the holotype of Celaenorhinus [sic!] phaeomelas Hübner, [1829] (type locality in Brazil) in MFNB reveals that it is sister to Mnasinous (Methionopsis) modestus (Godman, 1901) (type locality Mexico (Gue, Ver, Tab), Guatemala, Honduras, Panama, and South America to Brazil), the type species of the subgenus Methionopsis Godman, 1901, in the subtribe Falgina Grishin, 2019, and therefore does not belong to Papias Godman, 1900 (type species Pamphila integra Mabille, 1891), in the subtribe Moncina A. Warren, 2008, where it is currently placed (Fig. 67 magenta). The specimen we sequenced (Fig. 68) is indeed the holotype: it agrees with the original description and is a close match to the original illustrations of C. phaeomelas, is from the Herrich-Schäffer collection that included a number of Hübner types (now in MFNB), is labeled as “Hb. Orign.”, and, curiously, Oligoria lucifer (Hübner, [1829]) that is illustrated by Hübner ([1827]–[1829]) on the same plate with C. phaeomelas, also has a syntype extant in MFNB, originally from the Herrich-Schäffer collection. Because of the phylogenetic position of its holotype in the clade sister to the type species of the subgenus Methionopsis, we propose Mnasinous (Methionopsis) phaeomelas (Hübner, [1829]), comb. nov.
Fig. 67.

Papias monus (red), P. phainis (blue), P microsema (green), Mnasinous modestus (cyan), and M. phaeomelas (magenta). The figured syntype of P. microsema (olive, polytypic type series) from Panama is Cobalopsis valerius. The holotype of Celaenorhinus [sic!] phaeomelas is highlighted in yellow. Names shown are those proposed in this work.
Fig. 68.

Holotype of Mnasinous (Methionopsis) phaeomelas (Hübner, [1829]): a. dorsal (left), ventral (right), and labels (below), larger scale bar refers to specimens, smaller one is for labels; and b. its original illustrations (Hübner [1827]–[1829]) from the plate [100], figs. 581 (top) and 582 (bottom) rearranged and rotated, not to scale.
The type series of M. modestus included specimens from South America (Godman 1901) that are probably M. phaeomelas. However, Godman (1901: 599) writes: “we figure a male insect from Teapa” (Mexico: Tabasco), and, to define the identity of this name as suggested by the Recommendation 74B of the ICZN Code, N.V.G. hereby designates the specimen illustrated in Godman (1901), in the BMNH collection, that bears the following seven white labels, the first two round with a red circle, others rectangular: ( Type ), (Type | H. T. ) [ Teapa, | Tabasco. | March. H.H.S. ], [ ♂ ], [ Sp. figured. ], [ Godman-Salvin | Coll. 1913.—2.], [ B.C.A.Lep.Rhop. | Methionopsis | modestus, | Butl. ] as the lectotype of Methionopsis modestus Godman, 1901. This lectotype has left forewing with a tear at costa and genitalia expanded for inspection. Its abdomen was at some point detached to open genitalia and glued back. Finally, we note that M. phaeomelas (South American species) and M. modestus (North American species) are very close to each other genetically, e.g., their COI barcodes are basically identical (1 bp difference between the type specimens). However, the nuclear genome tree confidently partitions specimens into two clades (Fig. 67 magenta and cyan) and we keep M. phaeomelas and M. modestus as two distinct species pending further studies.
Papias microsema Godman, 1900, reinstated status
Currently treated as a junior subjective synonym of Celaenorhinus [sic!] phaeomelas Hübner, [1829] (type locality in Brazil), which in the section above we transferred from Papias Godman, 1900 (type species Pamphila integra Mabille, 1891) to the subgenus Methionopsis Godman, 1901 (type species Methionopsis modestus Godman, 1901), Papias microsema Godman, 1900 (type locality Mexico, (Tab), Costa Rica, Panama, Brazil (MT)) does not belong to Mnasinous (type species Mnasinous patage Godman, 1900) (Fig. 67) and therefore cannot be synonymous with Mnasinous (Methionopsis) phaeomelas. We demonstrate that the type series of P. microsema is polytypic. A syntype from Mexico: Tabasco, Teapa (NVG-21013E06 in CMNH) is indeed a distinct species of Papias (Fig. 67 green), and a syntype from Panama: Chiriqui (NVG-15035F01 in MFNB) whose ventral side is illustrated by Godman (1900) is Cobalopsis valerius (Möschler, 1879) (Fig. 67 olive). Although figured in the original illustration, this specimen was among the two that Godman commented about in the original description of P. microsema: “the spots … are quite distinct in … specimens … from Chiriqui, which we are unable … to dissect.” (Godman 1900). This phrase suggests that Godman considered these C. valerius specimens to be variations of his P. microsema rather than typical representatives. Godman defined his concept of P. microsema by genitalia: “P. microsema is exceedingly like P. phainis, but has differently formed genitalia.” Godman figured genitalia of a male from Mexico: Tabasco, Teapa, and this specimen belongs to Papias.
To define the identity of this name in a manner most consistent with the perceived intent of the original author, N.V.G. hereby designates the specimen with genitalia illustrated in Godman (1900), in the BMNH collection, that bears the following seven white labels, the first round with a red circle, others rectangular: ( Type ), [ Teapa, | Tabasco. | Feb. H.H.S. ], [ ♂ ], [ Sp. figured. ], [ 822 ], [ Godman-Salvin | Coll. 1914.—5.], [ B.C.A.Lep.Rhop. | Papias | microsema, | Godm. ] and a microslide with genitalia no. 822 pinned to the specimen with its labels, as the lectotype of Papias microsema Godman, 1900, stat. rest., which we reinstate as a species-level taxon. The lectotype has the anterior third of its left hindwing broken off and glued back. At this point, we are not able to determine the identity of Cobalus atramentarius Mabille, 1883 (type locality French Guiana: Cayenne), and for the lack of a better option, keep this name as a junior subjective synonym of M. phaeomelas while conducting research on this issue.
Papias unicolor (Hayward, 1938) and Papias monus Bell, 1942, reinstated status
Genomic sequencing of the holotype of Papias monus Bell, 1942 (type locality Guyana: Bartica) (Fig. 67 red) and a paralectotype of Papias phainis Godman, 1900 (type locality Mexico: Veracruz, Misantla) from Guatemala together with other specimens of this species from Central America (Fig. 67 blue), currently considered synonyms, reveals prominent genetic differentiation between them. E.g., their COI barcodes differ by 6.3% (42 bp), which is a distance typical of species from different subgenera. Moreover, a specimen from Brazil: São Paulo, NVG-21041F11, that we identified as Lerodea unicolor Hayward, 1938 (type locality in Paraguay) that is currently treated as another junior subjective synonym of P. phainis, is closely related, and yet separated in the genomic tree from compactly clustered Papias monus specimens collected across the species range (Fig. 67). Therefore, we propose to reinstate these taxa as species: Papias unicolor (Hayward, 1938), stat. rest. and Papias monus Bell, 1942, stat. rest. The two species are very close to each other genetically (only 0.3%, 2 bp COI barcode difference) and may be conspecific, but due to the genetic uniformity of P. monus across its wide range and the need to analyze more specimens of P. unicolor, including the holotype, we resort to the two-species treatment.
We also note that South American Papias projectus Bell, 1942 (type locality in Ecuador) is equally close to P. phainis: the same 0.3% (2 bp) COI barcode difference (Fig. 67). Likewise, additional work is needed to better understand the relationship between these two taxa, and they are kept as species pending further studies. Generally, we observe a marked genetic uniformity within species of Papias over large geographic distances (Fig. 67) and therefore even small, but consistent genetic differentiation may indicate speciation in this group of Hesperiidae.
Mnasilus guianae Lindsey, 1925 is confirmed as a junior subjective synonym of Papias amyrna (Mabille, 1891)
Placing a recently obtained genomic sequence of the holotype of Mnasilus guianae Lindsey, 1925 (type locality Guyana: Georgetown) (Fig. 69 magenta) in a phylogenetic context of specimens of Papias allubita (Butler, 1877) (type locality in Brazil: Para) and Papias amyrna (Mabille, 1891) (type locality Venezuela: Porto Cabello) that include a syntype of Pamphila amyrna Mabille, 1891 (NVG-15036F04) confirms our previous identification of specimens from Guyana and French Guiana as M. guianae and offers further support for Mnasilus guianae Lindsey, 1925 being treated as a junior subjective synonym of Papias amyrna (Mabille, 1891).
Fig. 69.

Papias amyrna (red, above, Mnasilus guianae holotype is labeled in magenta) and Papias allubita (blue, below).
Vidius pompeoides Grishin, new species
http://zoobank.org/5FA4FEDD-CCAD-4B2C-831A-46800F36442B
Fig. 70.

Holotype of Vidius pompeoides sp. n. dorsal (left) and ventral (right) views, data in text. Photographs by Bernard Hermier, © The Trustees of the Natural History Museum London and are made available under Creative Commons License 4.0 (https://creativecommons.org/licenses/by/4.0/).
Fig. 71.

Vidius pompeoides sp. n., (left) and Pompeius pompeius (right). iNaturalist observations 38437160, Suriname: Commewijne © Teri and 87229810, Brazil: DF, Brasília © Carlos A S Correia, respectively, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Definition and diagnosis. Inspection of a syntype of Cobalus catocala Herrich-Schäffer, 1869 (type locality not given) in MFNB reveals that it is not the species Evans identified as such. The syntype has pale postdiscal spots in cells RS-M1 and Sc+R1-RS on ventral hindwing aligned with each other, but in the specimens Evans identified as “Cobalopsis catocala” the spot in cell RS-M1 is closer aligned with the spot in cell M1-M2 and away from the spot in cell Sc+R1-RS (Evans 1955). We did not find an available name for Evans’ misidentification of C. catocala and therefore it is a new species that is described here. It keys to J.37.11 in Evans (1955) and is identified by the placement of spots on ventral hindwing as detailed above; hindwing brown, most spots are joined into two pale bands: subbasal and postdiscal, but the spot in cell Sc+R1-RS is offset to the base from the posdiscal band while remaining closer to it than to the subbasal band, and is separated from the subbasal band by a wide brown area; brown submarginal area frosted with pale scales, more extensive around the cells M1-M2 and M2-M3. Diagnosed by male genitalia with elongated nearly triangular harpe distally narrowing to a point; tegumen with uncus broad in dorsal view, approximately the same in width and length, uncus arms wide apart, hook-like. Differs from Pompeius pompeius (Latreille, [1824]) by the costal cell with a pale spot only at the base and no spot next to the postdiscal spot in cell Sc+R1-RS, and the pale spots defined sharper, with some darker overscaling inside the spots (Fig. 71); and from Vidius fraus (Godman, 1900) in having well-developed forewing hyaline spots and narrower ventral hindwing bands. This new species is assigned to Vidius Evans, 1955 (type species Narga vidius Mabille, 1891) due to wing pattern and genitalic similarities with V. fraus (Evans 1955), transferred from Cymaenes Scudder, 1872 to Vidius recently (Zhang et al. 2022b).
Type material. Holotype: ♂ deposited in the Natural History Museum, London, UK (BMNH), illustrated in Fig. 70, bears five labels: a round yellow ( 515 ), others rectangular, three white [ Sto. Paulo | Amazones | M de Mathan | 8bre.9bre.1879 ], [ PHOTO | AA ], [ R. Oberthür Coll. | Brit. Mus. 1931–136 ], and one red [ HOLOTYPE ♂ | Vidius pompeoides | Grishin ], and a small card pinned as a label with genitalia glued to it. Paratypes: 2♂♂ Brazil: Pará: Juruti (label data “Juhuty, Amazons”), April-1905, leg. M. de Mathan, in BMNH.
Type locality. Brazil: Amazonas, São Paulo de Olivença.
Etymology. By adding the suffix –oides, the adjectival name reflects superficial similarity of this species in its contrasting ventral wing pattern to frequently encountered and widely distributed Pompeius pompeius (Fig. 71 right) that has been confused with it (despite not being a close relative). For instance, the individual shown in Fig. 71 left was identified as P. pompeius on the iNaturalist website (2022) at the time of this writing.
Distribution. Known from the Amazonian region: Brazil (Amazonas and Pará) and Suriname.
Curva Grishin, new genus
http://zoobank.org/E623E221-BD77-4ADE-A01D-5C34E79B443B
Type species. Moeris hyagnis Godman, 1900.
Definition. Sister to Halotus Godman, 1900 (type species Halotus saxula Godman, 1900, which is a junior subjective synonym of Hesperia angellus Plötz, 1886) and is similar to it in the general shape of uncus and gnathos, but quite different in appearance and the shape of valva to be readily associated with Halotus (Fig. 72). Therefore, proposed here as a genus. Distinguished from its relatives by a unique shape of the valva with harpe concave at the distal margin and its dorsal margin smoothly folded over towards the aedeagus, as illustrated in the original descriptions of species we place in this genus (Godman 1900; Gaviria et al. 2018). In DNA, a combination of the following base pairs in nuclear genome is diagnostic: aly281.8.7:C389T, aly525.63.1:T1176C, aly2532.10.1:A2013G, aly2258.4.17:T108C, and aly276558.21.5:A255G.
Fig. 72.

Curva gen. n. (red, above) and Halotus (blue, below).
Etymology. The name is treated as a feminine noun in the nominative singular, given for the shape of the valva: Latin for curved, crooked, bent, or a fusion of Cur[ved] + [val]va, and, maybe an unintended Russian meaning for the difficulty of figuring these skippers out.
Species included. The type species (Fig. 73) and Psoralis darienensis Gaviria, Siewert, Mielke & Casagrande, 2018 (Fig. 74).
Fig. 73.

Curva hyagnis from Belize: Cayo District, Green Hills Butterfly Ranch. iNaturalist observation 37801765, color-corrected, rotated, and cropped, © Jan Meerman, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Fig. 74.

Curva darienensis comb. nov. from Panama: Darien, Cana, 1550 m, leg. G. B. Small [USNM]. a. ♂ NVG-22035F08, 10-Apr-1983; b. ♀ NVG-22035F07, 11-Apr-1983. Dorsal and ventral views are shown to the left and right from each letter.
Parent taxon. Subtribe Moncina A. Warren, 2008.
Comment. An alternative treatment could be to place this new genus as a subgenus of its sister Halotus, to which it is related phylogenetically (Fig. 72), but seems rather distinct from in genitalia, especially in the diagnostic shape of valva. Therefore, we treat the two as separate genera.
Nastra leuconoides (Lindsey, 1925), reinstated status
Genomic sequencing of its holotype reveals that Megistias leuconoides Lindsey, 1925 (type locality Brazil: Amazonas, Porto America) is not conspecific with Nastra leucone (Godman, 1900) (type locality in Guatemala) and instead is a close sister to Nastra subsordida (Mabille, 1891) (type locality in Honduras) (Fig. 75, Z chromosome tree). The COI barcodes of the two holotypes (M. leuconoides and N. subsordida) differ by 2% (13 bp). Therefore, we reinstate it as a species-level taxon Nastra leuconoides (Lindsey, 1925), stat. rest.
Fig. 75.

Nastra leuconoides (red), N. subsordida (blue), and N. leucone (magenta). Primary type labels are highlighted in green.
Rayia Grishin, new subgenus
http://zoobank.org/7EA724D4-4F7B-4809-A49E-DBD1DBCCECA5
Type species. Mastor perigenes Godman, 1900.
Definition. Currently included in the genus Nastra Evans, 1955 (type species Hesperia lherminier Latreille, [1824]), and is sister to and more distant from other species in the genus (Fig. 76). Due to its close relationship with Nastra and monotypic composition, it is proposed as a subgenus. Keys to J.24.6 in Evans (1955). Distinguished from its relatives by the following combination of characters: in addition to pale veins on the ventral hindwing, a pale ray from the wing base to near the outer margin along the anterior discal cell and vein M1 (Fig. 77); uncus with gnathos massive, nearly as broad as wide and tall; and valva nearly rectangular, harpe upturned, only slightly constricted in the middle, ending with a dorsally serrated narrow margin, distal margin convex, without prominent indentation. In DNA, a combination of the following base pairs is diagnostic in nuclear genome: aly276558.30.2:T42C, aly322.14.2: T418C, aly2379.9.2:A57G, aly1341.12.28:A8953C, and aly822.48.1: T354G; and in COI barcode: T85C, A241T, T457C, T499A, and A628T.
Fig. 76.

Nastra (Rayia) perigenes (magenta), N. fusca (red, neamathla its synonym, primary types labeled in red), and N. lherminier (blue).
Fig. 77.

Nastra (Rayia) perigenes from USA: Texas, Cameron Co. iNaturalist observation 8949490, brightened and cropped, © jamesgiroux, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Etymology. The name is a feminine noun in the nominative singular, given for the pale ray on the ventral hindwing of the type species.
Species included. Only the type species.
Parent taxon. Genus Nastra Evans, 1955.
Lerodea neamathla Skinner & R. Williams, 1923 is a junior subjective synonym of Nastra fusca (Grote & Robinson, 1867), reinstated status
Sequencing and comparative phylogenetic analysis of a syntype of Hesperia fusca Grote & Robinson, 1867 (type locality in USA: GA & FL), female in AMNH, currently a junior subjective synonym of Nastra lherminier (Latreille, [1824]) (type locality in USA: Carolina) places it in the clade with the holotype of Nastra neamathla (Skinner & R. Williams, 1923) (type locality in USA: Florida) (Fig. 76), and we conclude that the two type specimens are conspecific. Being a nearly completely brown female, this syntype is a challenge to identify as N. neamathla without genetic comparison. Nevertheless, closer inspection reveals traces of at least 2 pale small spots on each forewing in places characteristic of N. neamathla (Fig. 78) and almost never defined in females of N. lherminier. Because the name fusca has priority over neamathla leading to an undesirable name change, we looked for male syntypes (only one female syntype existed), hoping that, if found, at least one (out of three in existence) may be conspecific with N. lherminier as traditionally assumed. Nevertheless, it is also possible that all three male syntypes were conspecific with N. neamathla, because paler ventral hindwing veins characteristic of N. lherminier were not mentioned in the original description. Inspection of every Nastra specimen from Georgia and Florida in AMNH did not reveal any candidate syntypes, because all the specimens were either collected more recently, or, if the date was not given, there was nothing on their labels to associate them with the original description of H. fusca. We found a series of specimens in MFNB labeled as fusca, and although most of them were indeed N. lherminier, males from Georgia were collected by Morrison (and not Ridings or Linden as per description) and therefore are not syntypes. Hence, we assumed that the male syntypes were lost or are unrecognizable. First, to ensure nomenclatural stability and unambiguous identification of N. fusca, N.V.G. hereby designates its sole extant syntype, female in AMNH (Fig. 79), bearing the following five rectangular labels, the fourth one red and others white: [ Ga. ], [ No. 23113 | Grote & Robin ], [ Pamphila | fusca | type. G&R ], [ TYPE | No. | A. M. N. H. ], and [ Overlooked by | Beut. in his | various lists ] (Beut. is for William Beutenmüller) as the lectotype of Hesperia fusca Grote & Robinson, 1867. Second, this lectotype designation implies that Lerodea neamathla Skinner & R. Williams, 1923, syn. nov. is a junior subjective synonym of Nastra fusca (Grote & Robinson, 1867), stat. rest. On the one hand, the name change is not desirable, and the broader community of lepidopterists should decide whether to request ICZN to set the lectotype of N. fusca aside and designate a neotype conspecific with N. lherminier. On the other hand, the name fusca may be simpler than neamathla to remember.
Fig. 78.

Nastra fusca, stat. rest., formerly N. neamathla, USA: Florida, Alachua Co. iNaturalist observation 127295990 © Brian Ahern, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Fig. 79.

Lectotype of Nastra fusca (Grote & Robinson, 1867), stat. rest. with labels (not to scale), photos by Bernard Hermier.
Saturnus jaguar (Steinhauser, 2008), new combination
Parphorus jaguar Steinhauser, 2008 (type locality in Guyana, holotype NVG-15041A03 sequenced) (Fig. 80 olive) is not monophyletic with Parphorus Godman, 1900 (type species Phlebodes storax Mabille, 1891) (Fig. 80 red) and instead is very closely related to Saturnus metonidia (Schaus, 1902) (type locality in Brazil: Rio de Janeiro) within Saturnus Evans, 1955 (type species Papilio saturnus Fabricius, 1787) (Fig. 80 blue). Therefore, we propose Saturnus jaguar (Steinhauser, 2008), comb. nov. Despite close relationship between S. metonidia and S. jaguar as suggested by their 1.8% (12 bp) different COI barcodes, they are likely distinct species inhabiting different biogeographic zones.
Fig. 80.

Holotype of Parphorus hermieri sp. n. (inset shows magnified stigma) and the tree: Parphorus (red) and Saturnus (blue). Species discussed in the text are labeled in different colors.
Parphorus harpe (Steinhauser, 2008), new combination
Saturnus harpe Steinhauser, 2008 (type locality in Peru: Madre de Dios, holotype NVG-15038D04 sequenced) (Fig. 80 green) is not monophyletic with Saturnus Evans, 1955 (type species Papilio saturnus Fabricius, 1787) (Fig. 80 blue) and instead is siter to Parphorus Godman, 1900 (type species Phlebodes storax Mabille, 1891) (Fig. 80 red). Not willing to erect a monotypic genus for S. harpe due to genetic similarities: COI barcodes of its holotype and P. storax are 8.2% (54 bp) different, we propose to include it in Parphorus to form Parphorus harpe (Steinhauser, 2008), comb. nov. The tree reveals that Parphorus and Saturnus are closely related to each other, without long internal branches separating them, and P. harpe is a species that contributes to this lack of separation, originating early in evolution of the genus.
Parphorus hermieri Grishin, new species
http://zoobank.org/E5AD0DFC-354C-442F-8A56-C6F989C38C0D
(Fig. 80 part)
Definition and diagnosis. A species (Fig. 80 top and magenta in the tree) with unique for Parphorus Godman, 1900 (type species Phlebodes storax Mabille, 1891) (Fig. 80 red) predominantly yellow-orange wings, both dorsally and ventrally, reminiscent of Lento Evans, 1955 (type species Pamphila lento Mabille, 1878) (Fig. 80 orange) in color and pattern, but not closely related to Lento, originating within Parphorus close to its type species. Most similar to Lento kadeni Evans, 1955 (type locality not specified) in general appearance and wing patterns and therefore keys to I.3.4. in Evans (1955) but differs in being yellower rather than orange and in having brown-scaled areas narrower, e.g., ventral forewing with well-separated dark-brown spots near tornus instead of a continuous brown patch in L. kadeni; dorsal hindwing with yellow-orange ray along vein 1A+2A (only some orange scales, mostly hair-like, around this vein in L. kadeni but not a ray). Androconial patch triangular, filling the base of cell CuA1-CuA2, plus a small dash just below it and vein CuA2 (Fig. 80 top, inset).
Barcode sequence of the holotype: Sample NVG-21046D03, GenBank OP231469, 658 base pairs: AACTTTATATTTTATTTTTGGAATTTGAGCAGGAATATTAGGAACATCTTTAAGTTTATTAATTCGTACAGAATTAGGTAATCCTGGTTCTTTAATTGGGGATGATCAAATTTATAATACTATTGTAACAGCTCATGCATTTATCATAATTTTTTTTATAGTTATACCTATTATAATTGGAGGATTTGGTAATTGATTAGTTCCCTTAATATTAGGAGCTCCTGATATAGCTTTCCCTCGAATAAATAATATAAGATTTTGAATACTACCCCCTTCTTTAATACTATTAATTTCTAGAAGAATTGTAGAAAATGGTGCAGGAACTGGATGAACTGTTTACCCTCCTTTATCTTCAAATATTGCTCATCAGGGAGCTTCTGTTGATTTAGCAATTTTTTCTTTACACTTAGCAGGAATTTCTTCTATTTTAGGAGCTATTAATTTTATTACTACAATTATCAATATACGAATTAGAAATTTATCATTTGATCAAATACCTTTATTTGTTTGATCAGTAGGAATTACAGCACTTTTATTACTCTTATCTTTACCAGTGTTAGCTGGTGCTATTACTATACTTTTAACTGATCGAAATTTAAATACTTCATTTTTTGATCCTGCAGGAGGAGGAGATCCTATTTTATACCAACATTTATTT
Type material. Holotype: ♂ at the time of publication deposited in the McGuire Center for Lepidoptera and Biodiversity, Gainesville, FL, USA (MGCL), bears four rectangular printed labels: three white [ BRASIL: Rondonia | 3 km W of Candeias | on BR 364, 1 km S | on dirt road to Rio | Preto | 6 November 1995 | leg. G. T. Austin ], [ G.T. Austin colln. | MGCL Accession | # 2004–5 ], [ DNA sample ID: | NVG-21046D03 | c/o Nick V. Grishin ], and one red [ HOLOTYPE ♂ | Parphorus | hermieri Grishin ].
Paratype: ♂ Peru: Loreto, Castaña, −0.8037, −75.24, 150 m, 20-Oct-1993, leg. R. K. Robbins [MUSM].
Type locality. Brazil: Rondônia, 3 km west of Candeias do Jamari on interstate BR-364, then 1 km south on dirt road.
Etymology. The name honors Bernard Hermier (French Guiana), an expert on Neotropical Hesperiidae generously sharing his vast knowledge and time discussing challenging aspects of nomenclature and taxonomy, who encouraged to describe this species. Bernard’s kindness and generosity are unsurpassed and much valued by naturalists across the world. The name is a masculine noun in the genitive case.
Distribution. Known only from two specimens from Brazil: Rondônia and Peru: Loreto.
Comment. Due to superficial similarities of this new Parphorus species with Lento kadeni, we tentatively suggest Parphorus kadeni (Evans, 1955), comb. nov., reasoning (without solid evidence) that it fits better in Parphorus than in Lento. Sequencing of the P. kadeni holotype will test this taxonomic hypothesis.
Metiscus goth Grishin, 2022 confirmed as a species-level taxon by DNA
Proposed without DNA sequence data as a name for Evans’ concept of Enosis angularis infuscata (Plötz, 1882) (misidentification, Hesperia infuscata Plötz, 1882 is instead a junior subjective synonym of Mnaseas derasa derasa (Herrich-Schäffer, 1870) (Zhang et al. 2022b)), Metiscus goth Grishin, 2022 (type locality Costa Rica) is closely related to Metiscus angularis (Möschler, 1877) (type locality Suriname). Genomic sequencing of three M. goth paratypes and an additional specimen, and their comparison with M. angularis, including primary type specimens, reveals prominent genetic differentiation (Fig. 81) with Fst/Gmin statistics of 0.42/0.02, and their COI barcodes differ by 2.4% (16 bp). Therefore, we confirm M. goth as a species-level taxon and provide the COI barcode sequence of M. goth, identical in all four specimens (GenBank OP323110–OP323113): AACTTTATATTTTATTTTTGGTATTTGAGCAGGAATATTAGGAACTTCTTTAAGTTTATTAATTCGAACAGAATTGGGGAATCCTGGCTCTTTAATCGGAGATGATCAAATTTATAATACTATTGTAACTGCCCATGCTTTTATTATAATTTTTTTTATAGTTATACCTATTATAATTGGAGGATTTGGAAATTGACTAGTACCTTTAATATTAGGAGCCCCTGATATAGCTTTCCCACGAATAAATAATATAAGTTTTTGAATATTACCTCCTTCATTAATATTATTAATTTCAAGAAGAATTGTTGAAAATGGTGCAGGAACAGGATGAACAGTTTACCCCCCACTTTCTTCTAACATTGCCCATCAAGGATCTTCAGTAGATTTAGCAATTTTTTCTTTACATTTAGCAGGAATTTCCTCTATTTTAGGAGCTATTAATTTTATTACTACAATTATTAATATACGAATTAAAAGATTATCATTTGACCAAATACCTTTATTTGTTTGATCAGTAGGTATTACAGCTTTATTATTATTACTATCTTTACCTGTATTAGCTGGAGCTATTACCATACTTCTTACTGACCGAAATTTAAATACTTCATTTTTTGACCCAGCTGGTGGAGGAGATCCTATTTTATACCAACATTTATTT
Fig. 81.

Metiscus goth (red) and M. angularis (blue).
Panoquina ocola distipuncta Johnson & Matusik, 1988 is a subspecies of Panoquina lucas (Fabricius, 1793)
Genomic sequencing of a paratype of Panoquina ocola distipuncta Johnson & Matusik, 1988 (type locality in Dominican Republic) (Fig. 82 green) reveals that it does not belong to the clade with the neotype of Panoquina ocola ocola (W. H. Edwards, 1863) (type locality in USA: Florida) (Fig. 82 blue), but instead is in the clade with Panoquina lucas (Fabricius, 1793) (type locality “South American Islands”, meaning the West Indies) (Fig. 82 olive). This placement of P. ocola distipuncta within P. lucas is consistent with the wing patterns: both taxa possess a forewing discal cell hyaline spot typically lacking in P. ocola. Hence, P. ocola distipuncta is P. lucas, and we propose a new species-subspecies combination Panoquina lucas distipuncta Johnson & Matusik, 1988, comb. nov. As a result, P. ocola becomes monotypic.
Fig. 82.

Panoquina ocola (blue, above, neotype colored), P. lucas (olive, middle) with P. lucas distipuncta (green) as its subspecies, and Panoquina hecebolus (red, below) with Pamphila ortygia as its synonym (magenta).
Pamphila ortygia Möschler, 1883 is a junior subjective synonym of Panoquina hecebolus (Scudder, 1872)
Genomic sequencing of the holotype of Pamphila ortygia Möschler, 1883 (type locality in Suriname) currently treated as a junior subjective synonym of Panoquina ocola ocola (W. H. Edwards, 1863) (type locality in USA: Florida, neotype NVG-15097F07 sequenced) placed in the phylogenetic context of the three Panoquina Hemming, 1934 (type species Hesperia panoquin Scudder, 1863) species from the ocola group reveals that it is not associated with P. ocola (Fig. 82 blue) and instead belongs to Panoquina hecebolus (Scudder, 1872) (type locality Mexico: Oaxaca, Tehuantepec) (Fig. 82 red). Specimens of P. hecebolus do not show notable genetic differentiation across their range and we propose that Pamphila ortygia Möschler, 1883 is a junior subjective synonym of Panoquina hecebolus (Scudder, 1872), new synonym placement.
Zenis hemizona (Dyar, 1918) and Zenis janka Evans, 1955 are species distinct from Zenis jebus (Plötz, 1882)
The Z chromosome tree shows strong genetic differentiation between taxa of Zenis Godman, 1900 (type species Hesperia minos Latreille, [1824]) (Fig. 83) that have been treated as subspecies or synonyms of Hesperia jebus Plötz, 1882 (type locality in Brazil): Prenes hemizona Dyar, 1918 (type locality in Mexico) and Zenis jebus janka Evans, 1955 (type locality Panama: Bugaba). First, we agree with Evans’ (1955) identification of the nominotypical Zenis jebus, which is the only taxon that has four semi-equal short apical dashes on forewing, exactly as per original description. Second, compared to such a specimen from Brazil (NVG-18112F10), COI barcodes of Z. jebus hemizona holotype (NVG-18113F05) and Z. jebus janka from Costa Rica (NVG-7932) differ by 6.4% (42 bp) and 6.2% (41 bp), respectively. Therefore, we propose to reinstate Zenis hemizona (Dyar, 1918), stat. rest. and treat Zenis janka Evans, 1955, stat. nov. as species. Comparison of wing patterns of these species confirms that they typically can be distinguished by the number, relative size, and placement of forewing subapical hyaline spots. Furthermore, the tree reveals a new species-level taxon (Fig. 83 red) that is described below.
Fig. 83.

Species of Zenis: par sp. n. (red), hemizona (blue), janka (green), jebus (magenta), and minos (black).
Zenis par Grishin, new species
http://zoobank.org/319B5691-012A-42D5-8325-DF7D384333D9
Fig. 84.

Holotype of Zenis par sp. n. dorsal (left) and ventral (right) views, NVG-18112F09, data in text.
Fig. 85.

Zenis par sp. n., Ecuador: Napo, Archidona. iNaturalist observation 36717646. © Tom Horton, CC BY-NC 4.0 https://creativecommons.org/licenses/by-nc/4.0/
Definition and diagnosis. Sister to Zenis hemizona (Dyar, 1918) differing from it by 4.3% (28 bp) in the COI barcode, which, in the presence of wing pattern differences described below and likely stemming from notable nuclear genomic differentiation (Fig. 83) support species-level status of this taxon. This is the species that Evans (1955) identified (incorrectly) as Zenis jebus melaleuca. However, not all specimens misidentified by Evans as Z. j. melaleuca are this species: the female from “Espirito Santo” is Zenis jebus beckeri O. Mielke & Casagrande, 2002 (type locality in Brazil: Espírito Santo) that differs in the forewing apical spot pattern and has a white streak along ventral hindwing vein 1A+2A. This new species keys to O.3.2.(b). in Evans (1955). Distinguished from other Zenis species by nearly equal in length, strongly elongated apical hyaline spots in forewing cells R5-M1 (shorter spot) and M2-M3 (longer spot), dash-like well-developed hyaline spots in cells R2-R3, R3-R4, and R4-R5 adjoined to each other and separated by dark veins, the lack of a white streak along vein 1A+2A on hindwing ventral (Figs. 84, 85), and a combination of the following characters in the COI barcode: T38C, C235T, T478C, and T490C.
Barcode sequence of the holotype: Sample NVG-18112F09, GenBank OP231468, 658 base pairs: AACTTTATATTTTATTTTTGGAATTTGAGCAGGAATACTAGGTACTTCATTAAGTTTATTAATTCGAACAGAATTAGGAAATCCTGGTTCTTTAATCGGAGATGATCAAATTTATAACACTATTGTTACAGCACATGCTTTTATTATAATTTTTTTTATAGTTATACCTATTATAATTGGAGGATTTGGAAATTGATTAGTACCTTTAATATTAGGAGCCCCAGACATAGCTTTTCCTCGAATAAATAATATAAGATTTTGAATATTACCCCCTTCATTAACTTTATTAATTTCAAGAAGAATTGTAGAAAACGGTGCAGGAACAGGATGAACCGTTTACCCCCCTCTTTCTTCTAATATTGCTCATCAAGGTGCATCTGTTGATTTAGCAATTTTTTCTTTACATTTAGCAGGAATTTCATCAATTTTAGGAGCCATTAATTTTATTACTACAATTATTAATATACGAATTAAAAACTTATCATTTGACCAAATACCTTTATTTGTTTGATCAGTAGGTATTACAGCTTTATTATTACTTTTATCTCTTCCTGTTTTAGCTGGAGCTATTACTATATTATTAACTGATCGAAATTTAAATACATCTTTTTTTGATCCTGCTGGAGGAGGAGATCCTATTTTATACCAACATTTATTT
Type material. Holotype: ♂ in the National Museum of Natural History, Washington, DC, USA, to be deposited in the Museo de Historia Natural, Lima, Peru (MUSM), bears four rectangular printed labels: three white [ PERU: Cuzco 1194 m. | Quebrada Santa Isabel | Cosñipata Valley 5048 | 24-X-2016 Kinyon ], [ DNA sample ID: | NVG-18112F09 | c/o Nick V. Grishin ], [ USNMENT | {QR code} | 01531411 ], and one red [ HOLOTYPE ♂ | Zenis par | Grishin ]. Paratypes: 9♂♂, NVG-18092B10 from Ecuador: Morona-Santiago, Méndez, GPS −2.42, −78.20, 800 m, leg. J.-C. Petit, 16-Nov-2012, others (one specimen per date, all in MUSM) from Peru: Amazonas: Quebrada Chingaza, −5.367, −78.45, 500 m, 22-Sep-1999, leg. D. H. Ahrenholz; same data, but 24-Sep-1999, leg. G. Lamas; Cuzco: Cosñipata Valley: Quebrada Quitacalzón, −13.017, −71.50, 1050 m, 2-Apr-2015 & 1-Nov-2016, leg. S. Kinyon, and Quebrada Santa Isabel, −13.033, −71.517, 1200 m, 26-Jan-2020, leg G. Lamas; Madre de Dios: Puerto Maldonado, Lago Sandoval, [−12.583, −69.183], 200 m, 24-Oct-1990, leg. J. R. Macdonald; Tambopata Reserve, −12.833, −69.283, 300 m, 29-Oct-1991, leg. O. Mielke; Alto Río Madre de Dios, Albergue Amazonia, −12.867, −71.383, 500 m, 1-May-2015, leg. G. Lamas.
Type locality. Peru: Cuzco, Cosñipata Valley, Quebrada Santa Isabel, elevation 1194 m.
Etymology. The name, which is a Latin adjective for equal, stands for the two nearly equal in length dashes in forewing cells R5-M1 and M2-M3 that remind of an equal sign =.
Distribution. Currently confirmed by DNA from Ecuador and Peru, but likely present in Colombia and Amazonian region in Brazil.
Calpodes chocoensis (Salazar & Constantino, 2013), new combination
Placing genomic sequences of two specimens of Megaleas chocoensis Salazar & Constantino, 2013 (type locality Colombia: Valle del Cauca) (Fig. 86) in the context of Hesperiidae revealed a surprise. Instead of being close to Megaleas syrna (Godman & Salvin, 1879), the type species of Megaleas Godman, 1901, they were positioned deep within Calpodes Hübner, [1819] (type species Papilio ethlius Stoll, 1782) being a strongly supported sister to Calpodes severus (Mabille, 1895) (Fig. 87). Convergence with Megaleas in large yellow or orange forewing spots and divergence from the smaller, hyaline pale-yellow spots in Calpodes is the reason for this classification error. Genitalic morphology and the shape of forewing apical spots, however, support the relationship of M. chocoensis with Calpodes, and we propose to place it in this genus as Calpodes chocoensis (Salazar & Constantino, 2013), comb. nov. The COI barcode sequences of the two C. chocoensis specimens (Fig. 86) are identical (GenBank OP231470 & OP231471) and within 3% difference from some other Calpodes species, unambiguously indicating congeneric relationship: AACTTTATACTTTATTTTTGGTATTTGATCAGGAATATTAGGTACTTCATTAAGTTTATTAATTCGTACTGAATTAGGTAATCCTGGTTCTCTAATTGGAGATGATCAAATTTATAATACTATTGTTACAGCTCATGCTTTTATTATAATTTTTTTTATAGTTATACCTATTATAATTGGAGGATTCGGAAATTGATTAGTTCCTTTAATATTAGGTGCCCCTGATATGGCTTTCCCTCGAATAAATAATATAAGATTTTGAATACTTCCCCCTTCATTAACTTTATTAATTTCAAGAAGAATTGTAGAAAATGGTGCAGGAACAGGTTGAACAGTCTATCCCCCCCTTTCATCTAATATCGCCCACCAAGGATCATCAGTTGATTTAGCAATTTTTTCTTTACATTTAGCAGGAATTTCATCAATTTTAGGAGCTATTAATTTTATTACCACAATTATTAATATACGAATTAAAAATTTAATATTTGATCAAATACCATTATTTATTTGATCTGTAGGAATTACAGCATTATTATTATTATTATCTTTACCAGTTTTAGCAGGAGCTATTACTATATTACTTACTGATCGAAATTTAAATACATCTTTTTTTGACCCTGCAGGAGGAGGTGATCCTATTTTATACCAACATTTATTT
Fig. 86.

Sequenced specimens of Calpodes chocoensis from Colombia: Valle del Cauca, ~20 mi NW of Cali. a. ♂ NVG-21109C02, b. ♀ NVG-21109C03. Dorsal and ventral views are shown to the left and right from the letter. © Pierre Boyer.
Fig. 87.

Megaleas chocoensis (red) belongs to Calpodes (blue).
Carystus (Argon) argus Möschler, 1879, reinstated status, and Argentina as a likely type locality of Hesperia lota Hewitson, 1877
Genomic sequencing of the lectotype of Hesperia lota Hewitson, 1877 (type locality not specified), currently in the genus Carystus Hübner, [1819] (type species Papilio jolus Stoll, 1782), and the holotype of Carystus argus Möschler, 1879 (type locality in Colombia), currently a junior subjective synonym of the former, together with specimens from other localities reveals strong genetic differentiation between them (Fig. 88, Z chromosome tree). Carystus (Argon) lota is closely grouped with a specimen from Argentina, suggesting that the type locality of C. lota is in southeastern South America, possibly in Argentina. Due to this locality, we leave Pamphila cerymicoides Burmeister, 1878 (type locality Argentina: Misiones) as a junior subjective synonym of C. lota. All other specimens we sequenced (from Costa Rica, Panama, Colombia, and Dominican Republic) are in a different clade. COI barcodes of the two taxa differ by 2% (13 bp between their primary type specimens) and their Fst/Gmin statistics are 0.52/0.008. Therefore, we reinstate Carystus (Argon) argus Möschler, 1879, stat. rest. as a species. This resurrection from synonymy brings back the argus in the subtribe Carystina Mabille, 1878, and care should be taken not to confuse it with Oligoria (Cobaloides) argus Hayward, 1939 (type locality in Paraguay), a species from the subtribe Hesperiina that looks superficially similar to C. (A.) argus in wing shape, coloration, and the black-dotted pattern of ventral hindwing. A mnemonic to remember which argus is which may be that ‘O’ in Oligoria stands for a more rounded, arc-like arrangement of black hindwing dots: ‘O for round’; and ‘C’ in Carystus stands for a cluster of dots, i.e., several dots in the middle of the wing that do not form a long and smooth arc: ‘C for cluster’.
Fig. 88.

Carystus argus (red, above) and Carystus lota (blue, below). Primary type specimens are labeled in corresponding colors
Lycas devanes (Herrich-Schäffer, 1869), reinstated status, with the type locality in South America
Genomic sequencing of a syntype of Goniloba devanes Herrich-Schäffer, 1869 (type locality not specified), currently a junior subjective synonym of Lycas argentea (Hewitson, 1866) (type locality in Guatemala) (1955) places it in the clade of exclusively South American specimens that is separate from the clade with North American specimens including one from Guatemala (Fig. 89). The two clades show strong genetic differentiation with Fst of 0.66 and undetectable gene exchange between them. COI barcodes differ by 3.8% (25 bp) between the specimens from Guatemala (NVG-18111A12) and Argentina (NVG-17109H07) and by 3.6% (24 bp) between NVG-18111A12 and a specimen from Trinidad (NVG-18111B03). Therefore, the North American (i.e., L. argentea) and South American clades represent two distinct species. First, to ensure nomenclatural stability and unambiguous identification of G. devanes, N.V.G. hereby designates its sole syntype in MFNB bearing the following five rectangular labels, the first one red and others white: [ Typus ], [ gonil. devanes | m ], [ Coll. | Staudinger ], [ {QR code} http://coll.mfn-berlin.de/u/ | 449f7e ], and [ DNA sample ID: | NVG-15036C11 | c/o Nick V. Grishin ] as the lectotype of Goniloba devanes Herrich-Schäffer, 1869. The lectotype is missing the right antenna and scales are partly rubbed off the right forewing apex. It is likely that the second label is in Herrich-Schäffer’s handwriting and ‘m’ stands for ‘mihi’, which is Latin for ‘of me’, placed after a species name as an attribution of the new species to the writer. This notation was frequently used more than a century ago: ‘m’ or ‘mihi’ instead of the author’s name directly, as done nowadays. This ‘m’ further suggests that the label was written by Herrich-Schäffer and offers additional evidence of authenticity of this specimen as a syntype. Second, because the lectotype designated herein is in the clade consisting of South American specimens, we suggest that the type locality of G. devanes is in South America. Third, the two clades represent two distinct species, and therefore we propose species-level status for Lycas devanes (Herrich-Schäffer, 1869), stat. rest.
Fig. 89.

Lycas devanes (red, above) and L. argentea (blue, below). Name references are in corresponding colors.
Supplementary Material
ACKNOWLEDGMENTS
We acknowledge Ping Chen and Ming Tang for excellent technical assistance. We are grateful to David Grimaldi and Courtney Richenbacher (AMNH: American Museum of Natural History, New York, NY, USA), Blanca Huertas, David Lees, and Geoff Martin (BMNH: Natural History Museum, London, UK), Jonathan P. Pelham (BMUW: Burke Museum of Natural History and Culture, Seattle, WA, USA), Vince Lee and the late Norm Penny (CAS: California Academy of Sciences, San Francisco, CA, USA), Jim Fetzner, Bob Androw, Vanessa Verdecia, Cat Giles, and the late John Rawlins (CMNH: Carnegie Museum of Natural History, Pittsburgh, PA, USA), Chris Schmidt and Christi Jaeger (CNC: Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa, Ontario, Canada), Chuck Harp and the late Boris Kondratieff (CSUC: Colorado State University Collection, Fort Collins, CO, USA), Jason Dombroskie (CUIC: Cornell University Insect Collection, Ithaca, NY, USA), Crystal Maier and Rebekah Baquiran (FMNH: Field Museum of Natural History, Chicago, IL, USA), Weiping Xie (LACM: Los Angeles County Museum of Natural History, Los Angeles, CA, USA), Théo Léger, Wolfram Mey, and Viola Richter (MFNB: Museum für Naturkunde, Berlin, Germany), Andrei Sourakov, Andrew D. Warren, Debbie Matthews-Lott, and Keith R. Willmott (MGCL: McGuire Center for Lepidoptera and Biodiversity, Gainesville, FL, USA), Matthias Nuss and Manuela Bartel (MTD: Museum für Tierkunde, Dresden, Germany), Rob de Vos (RMNH: Naturalis Biodiversity Center, Leiden, Netherlands), Martin Wiemers and Christian Kutzscher (SDEI: Senckenberg Deutsches Entomologisches Institut, Müncheberg, Germany), Wolfgang A. Nässig (SMF: Natural History Museum, Frankfurt, Germany), Edward G. Riley, Karen Wright, and John Oswald (TAMU: Texas A&M University Insect Collection, College Station, TX, USA), Alex Wild (TMMC: University of Texas Biodiversity Center, Austin, TX, USA), Jeff Smith and Lynn Kimsey (UCDC: Bohart Museum of Entomology, University of California, Davis, CA, USA), Robert K. Robbins, John M. Burns, and Brian Harris (USNM: National Museum of Natural History, Smithsonian Institution, Washington, DC, USA), Ernst Brockmann (ZfBS: Zentrum fur Biodokumentation des Saarlandes, Schiffweiler, Germany), and Axel Hausmann, Andreas Segerer, and Ulf Buchsbaum (ZSMC: Zoologische Staatssammlung München, Germany), for granting access to the collections under their care, sampling specimens, and stimulating discussions; to Ernst Brockmann, Jim P. Brock, Bill R. Dempwolf, Bill Gendron, Howard Grisham, Bernard Hermier, Kiyoshi Maruyama, John Pasko, Harry Pavulaan, Steve M. Spomer, and Mark Walker for specimens and leg samples, to Christian Groneau and Bernard Hermier for photographs, to John V. Calhoun, Bernard Hermier, and Jonathan Pelham for critical reviews of the manuscript and many helpful suggestions and corrections. Evi Buckner-Opler assisted by providing emotional and logistic support and helped to collect specimens. We are indebted to California Department of Fish and Game for collecting permit SC13645, Texas Parks and Wildlife Department (Natural Resources Program Director David H. Riskind) for the research permit 08–02Rev, to U. S. National Park Service for the research permits: Big Bend (Raymond Skiles) for BIBE-2004-SCI-0011 and Yellowstone (Erik Oberg and Annie Carlson) for YELL-2017-SCI-7076, and to the National Environment & Planning Agency of Jamaica for the permission to collect specimens. Please note that photographs from iNaturalist (2022) reproduced in this work and photographs © The Trustees of the Natural History Museum, London are made available under Creative Commons License 4.0 (https://creativecommons.org/licenses/by/4.0/), which means in particular that when using the images you must give appropriate credit and provide a link to the license. We acknowledge the Texas Advanced Computing Center (TACC) at The University of Texas at Austin for providing HPC resources. The study has been supported in part by grants (to N.V.G.) from the National Institutes of Health GM127390 and the Welch Foundation I-1505.
Footnotes
ZooBank registration: http://zoobank.org/4EBE18FC-3018-49F4-8E2D-C6019918FF33
LITERATURE CITED
- Calhoun JV 2018. John Abbot, Jacob Hübner and Oreas helicta (Nymphalidae: Satyrinae). News of the Lepidopterists’ Society 60(4): 159–163. [Google Scholar]
- Cong Q, Shen J, Borek D, Robbins RK, Otwinowski Z, and Grishin NV. 2016. Complete genomes of hairstreak butterflies, their speciation, and nucleo-mitochondrial incongruence. Scientific Reports 6: 24863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cong Q, Shen J, Zhang J, Li W, Kinch LN, Calhoun JV, Warren AD, and Grishin NV. 2021. Genomics reveals the origins of historical specimens. Molecular Biology and Evolution 38(5): 2166–2176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cong Q, Zhang J, and Grishin NV. 2019a. Genomic determinants of speciation. bioRxiv BIORXIV/2019/837666. [Google Scholar]
- Cong Q, Zhang J, Shen J, Cao X, Brevignon C, and Grishin NV. 2020. Speciation in North American Junonia from a genomic perspective. Systematic Entomology 45(4): 803–837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cong Q, Zhang J, Shen J, and Grishin NV. 2019b. Fifty new genera of Hesperiidae (Lepidoptera). Insecta Mundi 0731: 1–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cramer P 1775–1780. De uitlandsche Kapellen voorkomende in de drie Waereld-Deelen Asia, Africa en America. Papillons exotiques des trois parties du monde l’Asie, l’Afrique et l’Amérique. Amsteldam: S.J. Baalde; Utrecht, Barthelemy Wild and J. Van Schoonhoven & Comp. Volumes 1–4. [Google Scholar]
- Draudt MWK 1921–1924. B. Grypocera, breitköpfige Tagfalter. In: Seitz A (Ed.). Die Gross-Schmetterlinge der Erde. Stuttgart: Alfred Kernen. 5: 833–1011, 1046–1139, pls 113B, 160–193. [Google Scholar]
- Eliot JN 1973. The higher classification of the Lycaenidae (Lepidoptera): a tentative Arrangement. Bulletin of the British Museum of Natural History (Entomology) 28(6): 373–505. [Google Scholar]
- Elwes HJ, and Edwards J. 1897. A revision of the Oriental Hesperiidae. Transactions of the Zoological Society of London 14(4): 101–324. [Google Scholar]
- Emmel JF, Emmel TC, and Mattoon SO. 2012. A new species of Cercyonis (Lepidoptera: Satyridae) from Northern California. Bulletin of the Allyn Museum 163: 1–12. [Google Scholar]
- Evans WH 1949. A catalogue of the Hesperiidae from Europe, Asia, and Australia in the British Museum (Natural History). London: The Trustees of the British Museum (Natural History). xix + 502 pp., 53 pls. [Google Scholar]
- Evans WH 1953. A catalogue of the American Hesperiidae indicating the classification and nomenclature adopted in the British Museum (Natural History). Part III. Pyrginae. Section 2. London: The Trustees of the British Museum (Natural History). v + 246 pp., pls. 26–53. [Google Scholar]
- Evans WH 1955. A catalogue of the American Hesperiidae indicating the classification and nomenclature adopted in the British Museum (Natural History). Part IV. Hesperiinae and Megathyminae. London: The Trustees of the British Museum (Natural History). v + 499 pp., pls. 54–88. [Google Scholar]
- Gaviria FG, Siewert RR, Mielke OHH, and Casagrade MM. 2018. A new species of Psoralis Mabille, 1904 from Panama (Lepidoptera, Hesperiidae, Hesperiinae, Moncini). Tropical Lepidoptera Research 28(1): 32–34. [Google Scholar]
- Godman FD 1900. Biologia Centrali-Americana. Insecta. Lepidoptera-Rhopalocera. London: Dulau & Co., Bernard Quaritch. 2(160): 557–588, pls. 99–100. [Google Scholar]
- Godman FD 1901. Biologia Centrali-Americana. Insecta. Lepidoptera-Rhopalocera. London: Dulau & Co., Bernard Quaritch. (162): 597–620, pls. 101–102. [Google Scholar]
- Haensch R 1909. 3. Familie: Danaidae. In: Seitz A (Ed.). Die Gross-Schmetterlinge der Erde. Stuttgart: Alfred Kernen. 5: 113–160. [Google Scholar]
- Hebert PD, Cywinska A, Ball SL, and deWaard JR. 2003. Biological identifications through DNA barcodes. Proceedings of the Royal Society B: Biological Sciences 270(1512): 313–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hemming AF 1937. Hübner. A bibliographical and systematic account of the entomological works of Jacob Hübner and of the supplements thereto by Carl Geyer, Gottfried Franz von Fröhlich and Gottlieb August Wilhelm Herrich-Schäffer. London: Royal Entomological Society. 1: xxxiv + 605 pp., frontisp.; 2: ix + [1] + 270 pp. [Google Scholar]
- Hübner J [1825]. Sammlung exotischer Schmetterlinge. Augsburg: Jacob Hübner. 2: pl. [120]. [Google Scholar]
- Hübner J [1827]–[1829]. Zuträge zur Sammlung exotischer Schmettlinge, bestehend in Bekundigung einzelner Fliegmuster neuer oder rarer nichteuropäischer Gattungen. Augsburg: Jacob Hübner. 3: 1–48. [Google Scholar]
- ICZN [International Commission on Zoological Nomenclature]. 1999. International code of zoological nomenclature. Fourth edition. London: International Trust for Zoological Nomenclature. xxx + 306 pp. [Google Scholar]
- iNaturalist. 2022. Available from https://www.inaturalist.org.
- Lamas G 2004. Checklist: Part 4A. Hesperioidea - Papilionoidea. In: Heppner JB (Ed.), Atlas of Neotropical Lepidoptera. Volume 5A. Gainesville: Association for Tropical Lepidoptera / Scientific Publishers. 439 pp. [Google Scholar]
- Li W, Cong Q, Shen J, Zhang J, Hallwachs W, Janzen DH, and Grishin NV. 2019. Genomes of skipper butterflies reveal extensive convergence of wing patterns. Proceedings of the National Academy of Sciences of the United States of America 116(13): 6232–6237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lukhtanov VA, Sourakov A, and Zakharov E. 2016. DNA barcodes as a tool in biodiversity research: testing pre-existing taxonomic hypotheses in Delphic Apollo butterflies (Lepidoptera, Papilionidae). Systematics and Biodiversity 14: 599–613. [Google Scholar]
- Mérit X, Manil L, Vila R, and Wiemers M. 2017. Inter-island differentiation of Leptotes (Cyclyrius) webbianus (Brullé, 1839) in the Canary Islands (Spain), with description of two new subspecies from La Palma and Gran Canaria (Lepidoptera: Lycaenidae). SHILAP-Revista de Lepidopterologia 45(179): 513–520. [Google Scholar]
- Mielke OHH 1975. Sobre algumas espécies de Staphylus Godman & Salvin (Lepidoptera: Hesperiidae). Acta biologica paranaense (Curitiba) 4(1/2): 25–34. [Google Scholar]
- Mielke OHH 2004. Hesperiidea, pp. 25–86. In: Lamas G (Ed.). Checklist: Part 4A. Hesperioidea - Papilionoidea. In: Heppner JB (Ed.), Atlas of Neotropical Lepidoptera. Volume 5A. Gainesville: Association for Tropical Lepidoptera / Scientific Publishers. [Google Scholar]
- Mielke OHH 2005. Catalogue of the American Hesperioidea: Hesperiidae (Lepidoptera). Curitiba: Sociedade Brasileira de Zoologia. 1: xiii + 125 pp.; 2: [ii] + 127–410; 3: [ii] + 411–771; 4: [ii] + 773–1055; 5: [ii] + 1057–1383; 6: [ii] + 1385–1536. [Google Scholar]
- Miller LD 1976. Revision of the Euptychiini (Satyridae) 3. Megisto Hübner. Bulletin of the Allyn Museum 33: 1–23. [Google Scholar]
- Pelham JP 2008. Catalogue of the Butterflies of the United States and Canada. Journal of Research on the Lepidoptera 40: 1–658. [Google Scholar]
- Pelham JP 2022. Catalogue of the Butterflies of the United States and Canada. Revised 2 February 2022. <http://www.butterfliesofamerica.com/US-Can-Cat.htm> Accessed 25 August 2022.
- Plötz C 1886. Nachtrag und Berichtigungen zu den Hesperiinen. Stettiner entomologische Zeitung 47(1/3): 83–117. [Google Scholar]
- Reuss T [1926]. Systematischer Überblick der Dryadinae T. Rss. mit einigen Neubeschreibungen (Lep. Rhopal.). Deutsche Entomologische Zeitschrift 1926(1): 65–70. [Google Scholar]
- Ribbe C 1889. Beiträge zur Lepidopteren-Faun von Gross-Ceram. Deutsche entomologische Zeitschrift Iris 2(2): 187–265. [Google Scholar]
- Robbins RK, Cong Q, Zhang J, Shen J, Busby RC, Faynel C, Duarte M, Martins ARP, Prieto C, Lamas G, and Grishin NV. 2022. Genomics-based higher classification of the species-rich hairstreaks (Lepidoptera: Lycaenidae: Eumaeini). Systematic Entomology 47(3): 445–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rothschild W, and Jordan K. 1903. A Revision of the Lepidopterous Family Sphingidae, Part 1. London and Aylesbury: Hazell, Watson & Viney, Limited. 813 pp. [Google Scholar]
- Scott JA 1994. Biology and systematics of Phyciodes (Phyciodes). Papilio (n.s.) (7): 1–120. [Google Scholar]
- Scott JA 1998. Phyciodes (Phyciodes): new discoveries, new subspecies, and convergence. Papilio (n.s.) 10: 1–42. [Google Scholar]
- Scott JA 2006. Phyciodes (Phyciodes): more progress. Papilio (n.s.) 13: 1–34. [Google Scholar]
- Scott JA 2014. The identity of Coenonympha brenda W. Edwards 1870 (Nymphalidae). Papilio (n.s.) 22: 8–16. [Google Scholar]
- Shen J, Cong Q, Borek D, Otwinowski Z, and Grishin NV. 2017. Complete genome of Achalarus lyciades, the first representative of the Eudaminae subfamily of skippers. Current Genomics 18(4): 366–374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen J, Cong Q, Kinch LN, Borek D, Otwinowski Z, and Grishin NV. 2016. Complete genome of Pieris rapae, a resilient alien, a cabbage pest, and a source of anti-cancer proteins. F1000Research 5: 2631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warren AD 2005. Butterflies of Oregon: Their Taxonomy, Distribution, and Biology. Lepidoptera of North America 6: 1–408. [Google Scholar]
- Zhang J, Cong Q, Burns JM, and Grishin NV. 2022a. Checking the checkered taxonomy of Plötz’s checkered skippers (Hesperiidae: Pyrgini). The Taxonomic Report of the International Lepidoptera Survey 10(5): 1–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Cong Q, Shen J, Brockmann E, and Grishin NV. 2019a. Genomes reveal drastic and recurrent phenotypic divergence in firetip skipper butterflies (Hesperiidae: Pyrrhopyginae). Proceedings of the Royal Society B: Biological Sciences 286(1903): 20190609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Cong Q, Shen J, and Grishin NV. 2022b. Taxonomic changes suggested by the genomic analysis of Hesperiidae (Lepidoptera). Insecta Mundi 0921: 1–135. [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Cong Q, Shen J, Opler PA, and Grishin NV. 2019b. Changes to North American butterfly names. The Taxonomic Report of the International Lepidoptera Survey 8(2): 1–11. [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Cong Q, Shen J, Opler PA, and Grishin NV. 2019c. Genomics of a complete butterfly continent. bioRxiv BIORXIV/2019/829887. [Google Scholar]
- Zhang J, Cong Q, Shen J, Opler PA, and Grishin NV. 2020. Genomic evidence suggests further changes of butterfly names. The Taxonomic Report of the International Lepidoptera Survey 8(7): 1–40. [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Cong Q, Shen J, Opler PA, and Grishin NV. 2021. Genomics-guided refinement of butterfly taxonomy. The Taxonomic Report of the International Lepidoptera Survey 9(3): 1–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Cong Q, Shen J, Song L, and Grishin NV. 2022c. Genomic DNA sequencing reveals two new North American species of Staphylus (Hesperiidae: Pyrginae: Carcharodini). The Taxonomic Report of the International Lepidoptera Survey 10(4): 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Shen J, Cong Q, and Grishin NV. 2019d. Genomic analysis of the tribe Emesidini (Lepidoptera: Riodinidae). Zootaxa 4668(4): 475–488. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.