

Abstract
A comprehensive analysis of the structure of neuronal nitric oxide synthase (nNOS; EC 1.14.13.39) mRNA species revealed NOS1 to be the most structurally diverse human gene described to date in terms of promoter usage. Nine unique exon 1 variants are variously used for transcript initiation in diverse tissues, and each is expressed from a unique 5′-flanking region. The dependence on unique genomic regions to control transcription initiation in a cell-specific fashion burdens the transcripts with complex 5′-mRNA leader sequences. Elaborate splicing patterns that involve alternatively spliced leader exons and exon skipping have been superimposed on this diversity. Highly structured nNOS mRNA 5′-untranslated regions, which have profound effects on translation both in vitro and in cells, contain cis RNA elements that modulate translational efficiency in response to changes in cellular phenotype.
Neuronal nitric oxide synthase (nNOS; EC 1.14.13.39) plays protean roles in a spectrum of physiological and pathological processes (1–3). To gain insight into the molecular basis of nNOS regulation it is necessary to have a comprehensive understanding of how the gene is regulated. Given that expression patterns of the nNOS mRNA and protein are complex with respect to the developmental cascade and cell lineages involved, it is plausible that the regulatory mechanisms that are operative may provide a unique model within the neurosciences. What are the basic principles that determine when, where, and how much nNOS enzyme is expressed in such diverse tissues?
Genes that play key roles in complex processes are often regulated at multiple levels. One such regulatory mechanism is alternative usage of multiple promoters. For instance, at least five independent promoters direct transcription of the human dystrophin gene in a cell-specific and developmentally controlled fashion (4). Other examples include the genes for c-Myc (5, 6), platelet-derived growth factor β receptor (7), and aromatase cytochrome P450 (8). Clearly, variation in the location of RNA polymerase II transcription initiation on genomic DNA affects mRNA sequence and structure. In this regard, alternative promoter usage may affect the coding region, changing the structure of the encoded protein, or in other examples, the 5′ untranslated region (UTR) is solely affected, resulting in differences in the sequence of 5′-leaders that have a profound impact on RNA biology. Mechanisms by which 5′-leaders modulate the fate of RNA include, but are not limited to, regulation of cap-dependent and -independent translation (9, 10), RNA stability (11), and RNA polymerase II processivity (12). Arguably, most attention has focused on cap-dependent translation, revealing that 5′-UTR structure determines the efficiency of translation, in part, through cis effects at the primary (e.g., upstream short ORFs affecting ribosome scanning), secondary (e.g., hairpins and stem-loop structures), or tertiary (e.g., pseudoknots) levels. Such effects often involve the contribution of RNA-binding proteins (e.g., IRP-1) (13–15). It is clear that structural diversity in 5′-UTR leader sequences may affect gene expression through this pathway. In this regard, it is noteworthy that discordant expression of nNOS mRNA and protein has been observed. The nNOS mRNA transcript is expressed very early during development but nNOS protein is evident only later, hinting at the existence of a translational regulatory mechanism (16). Last, characterization of the structural organization of the human nNOS gene implied the existence of a large primary transcript in neurons (160 kb) (17, 18). It could be argued that it would be inefficient to rely solely upon transcription initiation for the control of nNOS gene expression, given this extreme size.
On the basis of this background we performed a comprehensive analysis of the structure of mRNA species derived from the human nNOS gene. We report the cloning of nine independent variants of nNOS exon 1, their structural organization in the human genome, and their tissue-specific expression. Complex splicing patterns involving alternatively spliced 5′-UTR exons are described, as are exonic deletions involving exon 2. Importantly, we demonstrate that this diversity at the 5′ end of nNOS mRNA transcripts affects translational efficiency, and we posit its importance in the tissue- and developmental stage-specific expression of the gene.
Methods
Rapid Amplification of 5′ cDNA Ends (5′-RACE).
Total cellular RNA (1 μg) derived from various normal human tissues (CLONTECH) was reverse transcribed with a gene-specific antisense primer (P1, exon 2, 5′-CCA TGG TAA CTA GCT TCC-3′), as described (17). First round PCR was performed for 35 cycles with primer annealing at 55°C (generic sense primers 5′-GAC TCG AGT CGA CGA ATT CAA (T)17-3′, 2.5 pmol; 5′-GAC TCG AGT CGA CGA ATT CAA-3′, 25 pmol; gene-specific antisense primer P2, exon 2, 5′-TCT CTA AGG AAG TGA TGG TTG-3′, 25 pmol). A second round of amplification was performed for 35 cycles with primer annealing at 57°C (generic sense primer 5′-GAC TCG AGT CGA CGA ATT CAA-3′, 25 pmol; nested gene-specific antisense primer P3, exon 2, 5′-GGC TGT GTC TAG AAG TGA CG-3′, 25 pmol). PCR products were digested with EcoRI/XbaI, subcloned into pBluescript SK (−), and subjected to DNA sequence analysis.
Genomic Characterization.
P1 clones were isolated by PCR-based screening from a library of human diploid genomic DNA partially digested with Sau3A I in the vector pAD10SacBII (Genome Systems, St. Louis). Three overlapping P1 clones were mapped with the method of partial enzymatic digestion using EcoRI, MluI, SalI, and SmaI on a CHEF III DR mapping system (Bio-Rad). All exonic sequences and exon–intron boundaries were determined on both strands of genomic DNA with an ABI PRISM 377 automated DNA sequencer (Applied Biosystems, Perkin-Elmer Cetus).
Reverse Transcription (RT)-PCR.
First-strand cDNA was synthesized with total cellular RNA (5 μg) derived from varied normal human tissues (CLONTECH) by using random primers. The primers used for PCR amplifications to assess the tissue specificity of alternative exon 1 sequences and overall nNOS expression are described in supplemental data on the PNAS web site (www.pnas.org; Table 1). PCR amplifications were performed under well-characterized semiquantitative conditions in a total volume of 100 μl for 30 cycles. An 880-bp fragment of DNA encoding human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was amplified as a quantitative control (sense primer: 5′-ACA TCG CTC AGA CAC GAT GG-3′; antisense primer: 5′-GCT GTT GAA GTC AGA GGA GAC C-3′). PCR products were size-fractionated by agarose gel electrophoresis and downward transferred to GeneScreenPlus membranes (DuPont). Southern blots were hybridized with [γ-32P]ATP-labeled oligonucleotides positioned internally to flanking PCR primers.
RNase Protection Analysis.
[α-32P]CTP antisense riboprobes were synthesized by using 5′-RACE clone DNA templates and a T7 MAXIscript kit (Ambion, Austin, TX). For exon 1c, linearization at an internal StyI site resulted in a 318-nt probe complementary to 200 nt of exon 1c and 91 nt of exon 2. Linearization at an internal exon 1f ApoI site resulted in a 262-nt probe complementary to 144 nt of exon 1f and 91 nt of exon 2. Linearization of the exon 1g template at a vector-encoded HindIII site resulted in a 297-nt probe complementary to 151 nt of exon 1g and 91 nt of exon 2. Gel-purified probe (105 cpm) was hybridized for 16 hr at 42°C with yeast tRNA (50 μg) or human total cellular RNA (50 μg) (CLONTECH). RNase protection was performed with the RPA II kit (Ambion), as described (17).
In Situ Hybridization.
In situ hybridization was performed as previously described (19). Full-length antisense cRNA probes were transcribed using viral RNA polymerases (Promega) in the presence of α-[35S]thio-UTP (specific activity 1,200 Ci/mmol; Amersham; 1 Ci = 37 GBq) from linearized cDNA clones representing full-length exons 1a, 1c, 1f, and 1g. cRNA probes transcribed in the sense orientation were used as negative controls. Tissue collection was performed in accordance with Emory University guidelines and was approved by the Emory University Hospital Human Investigations Committee.
Translation Assays.
In vitro transcription of capped RNA was carried out with the Sp6 mMessage mMachine kit (Ambion) and multiple independent DNA preparations. The quality of in vitro synthesized, m7GpppG-capped RNA was assessed on 0.66 M formaldehyde/1% agarose gels. In vitro translation was performed in rabbit reticulocyte lysates (Promega). C2C12, NT2/D1, HeLa, and JEG-3 cells (American Type Culture Collection) were maintained in 60-mm dishes and transfected at 40–60% confluency with in vitro synthesized, quantified, capped RNA by using Lipofectin (Life Technologies). Three micrograms of RNA and 12 μl of Lipofectin were used for each 60-mm dish along with 5 μg of β-galactosidase RNA to control for RNA transfection efficiency. Differentiation of C2C12 cells was induced 60 hr before transfection, and ≥80% of cells were fused at the time of transfection. Cell extract was harvested at 6–8 hr post transfection. Multiple independent RNA preparations were used. Northern analysis using total cellular RNA harvested 5–7 hr post transfection and a random-primer-labeled luciferase ORF DNA probe was used to assess steady-state levels of transfected reporter RNA molecules. For DNA transient expression, C2C12 cells were infected with Sp6 recombinant vaccinia virus 30 min before transfection with 1 μg of reporter DNA constructs driven by an Sp6 promoter and 0.5 μg of pRSV-βgal, using 6 μl of Lipofectin per 60-mm dish. Cell extract was harvested at 30 hr post transfection. Alternatively, NT2/D1 and C2C12 cells were transfected with a mixture of 1 μg of reporter DNA constructs under the transcriptional control of simian virus 40 (SV40) promoter, 1 μg of pRSV-βgal plasmid, 2 μg of pBluescript plasmid, and 8 μl of Lipofectin per 60-mm dish. Cell extract was harvested at 48 hr post transfection. Luciferase activity was measured with a luminometer (Monolight 2010C, Analytical Luminescence Laboratory, Sparks, MD) and normalized for β-galactosidase activity and protein content.
Results
Cloning of Multiple Exon 1 Variants and Their Genomic Organization.
We used the 5′-RACE method and RNA isolated from different human tissues to characterize the structure of nNOS mRNA 5′ termini. The design of the RACE strategy allowed cloning of varied 5′ sequences upstream of exon 2 where translation of nNOS initiates (Fig. 1A). A total of 97 RACE clones were analyzed (Table 2 in supplemental data, www.pnas.org). All clones exhibited a common 3′ sequence that was identical to the exon 2 sequence of human nNOS. They differed, however, in the sequences upstream of the first nucleotide of exon 2. Nine distinct cDNA 5′ termini were identified, including the limited number we and others have previously reported (3, 17, 18, 20). Subsequent characterization of three P1 bacteriophage clones representing genomic regions upstream of exon 2 indicated that each of the nine unique 5′ termini was encoded by a single distinct exon that was spliced to the common exon 2 (Fig. 1B and supplemental data, www.pnas.org). These nine exon 1 variants, arising from alternative promoters from a single nNOS gene, were distributed over a region greater than 105 kb and designated as exons 1a, 1b, 1c, 1d, 1e, 1f, 1g, 1h, and 1i, on the basis of their 5′-to-3′ order in the human genome at 12q24.2 (17). All exonic sequences were confirmed on both strands of the genomic DNA and submitted to the GenBank databank (accession nos. AF049712-AF049720). All exon–intron boundaries conformed to the GT/AG rule (Fig. 1B). Since stop codons were evident in all three translation reading frames immediately upstream of the initiating AUG in exon 2 (17), all newly identified sequences represented nNOS mRNA diversity within the 5′-UTR and did not affect the encoded protein sequence.
Figure 1.

Human nNOS 5′-mRNA structure and cloning results. Nine distinct examples of exon 1 were isolated with 5′-RACE in 97 cDNA clones. (A) Exon 1 splices to a common exon 2 where translation initiates. Shaded area represents the coding region. The gene-specific antisense primers used in the 5′-RACE are depicted as arrowheads P1, P2, and P3. Diversity arising from downstream promoter activity would not be detected (32). (B) Nucleotide sequences of varied exon 1 examples and exon AS of the human nNOS. Uppercase letters, exonic sequences; lowercase letters, intronic sequences. GenBank accession nos. AF049712–AF049720.
Tissue-Specific Expression of Alternative Exon 1 Variants.
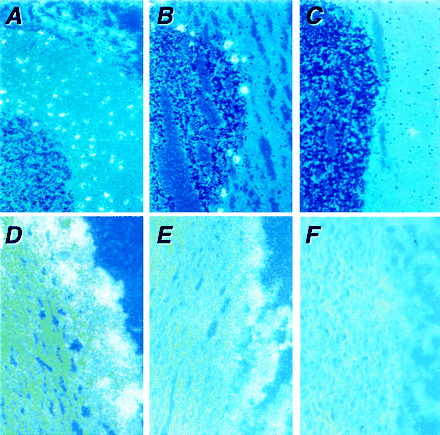
A semiquantitative RT-PCR approach, RNase protection analyses, and in situ hybridization were used to determine the tissue-specific expression of each human exon 1 in normal tissues. PCR amplification was performed using exon 1 sense and exon 2 or exon 3 antisense primers (Fig. 2). Consistent with findings from 5′-RACE, exon 1 sequences were expressed in a tissue-specific fashion compared with amplifications subtending common regions, such as exons 2/3 or 20/21 (Fig. 2A). Notably, exons 1a, 1b, and 1c were robustly expressed in skeletal muscle, whereas exons 1f, 1g, and 1i were relatively abundant in brain. Some tissues, such as testis, expressed all nine exon 1 variants. This finding contrasted with a limited repertoire of exon 1 usage in select tissues. For instance, exon 1g was prominently expressed in lymph node. Varied 5′-intron donor sites were used uncommonly by three exon 1 variants (exons 1b, 1e, 1i). The existence of a distinct set of alternative exon 1 variants implies diverse and exquisite regulation at the transcriptional level. RNase protection results corroborated the restricted tissue-specific expression of the varied exon 1 examples (supplemental data, www.pnas.org). For instance, exon 1c was enriched in skeletal muscle, whereas exons 1f and 1g were enriched in cerebellum. These assays used varied exon 1-specific cRNA probes each representing portions of exon 1 as well as exon 2. Protected fragments corresponded to unique exon 1/exon 2 species, common exon 2 regions, and/or portions of exon 1 if a nNOS mRNA transcript had undergone a 5′ alternative splicing event (i.e., exon AS). We also performed a comprehensive series of in situ [35S]thio-UTP-labeled antisense cRNA hybridizations with the varied exon 1-specific probes (supplemental data, www.pnas.org). These studies confirmed (i) that expression of nNOS exon 1 variants was cell restricted (e.g., exons 1f and 1g were differentially expressed in the cerebellar granular layer and Purkinje cells, whereas exon 1a was not expressed in cerebellum or aorta) and (ii) that a single cell type could express multiple nNOS exon 1 variants (e.g., exons 1f and 1g in the perivascular neurons of the aorta). Further studies will be necessary to characterize the transcription initiation start sites and regulatory mechanisms underlying these complex nNOS transcriptional processes. Interestingly, genomic regions encoding exon 1 variants enriched in skeletal muscle were clustered together in an upstream region, whereas those genomic regions encoding examples of exon 1 enriched in neuronal tissues were grouped in a common region 75 kb further downstream. This unusual arrangement of putative promoters on the genome indicates the existence of tissue-specific control mechanisms that serve to coordinate transcription initiation, elongation, and splicing of nNOS mRNA transcripts.
Figure 2.

Tissue-specific expression of various exon 1 examples and their splicing patterns assessed with semiquantitative RT-PCR. B, brain; S, skeletal muscle; K, kidney; A, adrenal gland; H, heart; T, testis; L, lung; N, lymph node. Locations of PCR primers are indicated for each blot at the far left. (A) PCR products derived from amplification between exon 1 examples and exon 2. Southern blots were probed with an oligonucleotide located in exon 2 common region and hybridized to PCR products both with (larger) and without (smaller) the exon AS insertion. The 553-bp product in 1b/2 represents exon 1bL (see B). PCR amplifications between exons 2 and 3 and exons 20 and 21 were used to assess overall nNOS expression. (B) Blots were reprobed with an oligonucleotide located within exon AS. PCR amplification between exons AS and 2 is also shown. (C) PCR products derived from amplification between exons 1a, 1c, 1e, and exon 3 and probed with an exon 3 oligonucleotide. Note that for exon 1e cassette deletion of exon 2 was a common event where a is exon 2 deletion, b is exon AS insertion plus exon 2 deletion, and c is exon 1eL with exon AS insertion plus exon 2 deletion. Shown in A–C are representative results from one of three experiments.
Complex Alternative Splicing Patterns.
Two intriguing features of nNOS primary RNA processing were also evident. First, an alternative splicing event was revealed. Interposed between varied exon 1 examples and the common exon 2 was an 89-nt exon (exon AS), the expression pattern of which exhibited exon 1 and tissue specificity (Fig. 2 A and B). For instance, the cassette insertion event was frequent in nNOS mRNA transcripts initiating with neuronal exon 1g, less frequent with neuronal exon 1f, and barely detected in exon 1d transcripts. Insertion of exon AS was not detected in nNOS mRNA transcripts expressed in lymph node tissue, indicating further that products of this splicing event were relatively enriched in certain tissues. An interesting genomic structural arrangement was observed for exon 1i. Exon AS was contiguous with the 3′ end of exon 1i, indicating that the same cis region could serve either as exon 1 or as a 3′-intronic splicing acceptor. Inclusion of exon AS markedly increased the potential number of different nNOS mRNA transcripts. Second, cassette deletion of the large (1145-nt) exon 2 was a common pattern in the expression of the human nNOS gene (Fig. 2C). This would lead to structural alterations in the nNOS protein because translation initiates in this exon. This deletion event appeared to be promoter- and tissue-specific. For instance, exon 2 was deleted in the vast majority of the exon 1e transcripts in skeletal muscle (Fig. 2 A and C). Deletion of exon 2 was originally described in an nNOSΔ/Δ mouse, wherein exon 2 was disrupted by homologous recombination (3, 21).
Effects of Distinct nNOS 5′-UTRs on Translation.
Taken together, the remarkable diversity at the 5′ terminus of nNOS mRNA transcripts is, to our knowledge, unprecedented in any reported mammalian gene. Not obvious is the biological relevance of this complexity. We posited that the dependence on unique genomic regions to control transcription initiation in a tissue-specific fashion burdened the transcript with complex 5′-mRNA leader sequences. Sequence inspection of representative examples of the various full-length 5′-UTRs defined by 5′-RACE, RT-PCR, and RNase protection highlighted the following features. First, leader sequences ranged in size from 489 to 1033 nt. This size is considerably larger than the average length of eukaryotic 5′-UTRs (50–100 nt) (13). Second, multiple upstream AUG codons were detected in all sequences indicating the existence of multiple short ORFs. Also, composition analysis revealed that some sequences were highly enriched in G+C content (e.g., 68%, 79%, and 70% of exons 1c, 1g, and 1h were G or C, respectively). Finally, secondary mRNA structure of the varied 5′-UTR nNOS sequences was modeled by using the GCG Wisconsin package (Fold/Squiggles subroutine, v 8.0) which indicated that all were likely to form complex and stable structures (Table 3 in supplemental data, www.pnas.org). These features are regarded as important determinants of translational efficiency (13–15). This suggested that the diversity at the 5′ end of nNOS mRNA transcripts could have important biological effects on the regulation of translation and possibly explain reported discrepancies between steady-state mRNA levels and protein expression (16).
We created reporter luciferase constructs to functionally assess the effects of nNOS 5′-UTR sequences on translational efficiency (Fig. 3A). Marked differences in reporter activity were evident when in vitro transcribed and capped RNAs were quantified and translated in rabbit reticulocyte lysates (Fig. 3B). The effects depended on which nNOS leader sequence was incorporated. Sequences corresponding to shared exon 2 5′-UTR regions repressed reporter translation more than 10-fold. Addition of exon 1 sequences resulted in further repression of translation in a sequence-specific fashion. For example, incorporation of exons 1a or 1b had minor additional effects compared with exon 2 alone, whereas the sequences from exons 1c, 1e, 1f, 1g, or 1h repressed translation a further 10- to 20-fold. Overall, exon 1 sequences that were enriched in neuronal tissues exerted a more robust repressive effect on translational efficiency than those enriched in skeletal muscle. These results indicate that the diverse nNOS 5′-UTR sequences markedly affect nNOS gene expression at the level of translation, at least in vitro.
Figure 3.

Effects of nNOS 5′-UTR sequences on translational efficiency. (A) Schematic diagram of the chimeric reporter constructs. Various nNOS 5′-UTR sequences were inserted between the Sp6 viral RNA polymerase promoter and the firefly luciferase reporter ORF. (B) In vitro transcribed and capped RNAs were quantified and translated in rabbit reticulocyte lysates. In these experiments vector luciferase activity averaged 4.2 ± 0.2 × 106 relative light units (RLU). (C) Effects of the various nNOS leader sequences on the translational efficiency of in vitro transcribed, capped, chimeric nNOS 5′-UTR/luciferase reporter RNAs that were transiently expressed in cultured cells. Activity profiles of nNOS 5′-UTR sequences in C2C12 cells [mouse skeletal muscle, nNOS(+)], NT2/D1 cells [human neuroblastoma, nNOS(+)], and HeLa cells [human squamous carcinoma, nNOS(−)] were normalized with β-galactosidase activity and protein content. In these experiments, vector luciferase activity averaged (5.6 ± 0.4) × 105 RLU for nondifferentiated C2C12 cells, (4.6 ± 0.9) × 105 RLU for NT2/D1 cells, and (5.1 ± 0.9) × 105 RLU for HeLa cells. (D) Translational efficiency of selected nNOS 5′-UTR sequences examined by using liposome-mediated DNA transfections. Expression of RNA molecules from Sp6 polymerase sequences employed infection of nondifferentiated C2C12 cells with recombinant vaccinia vectors expressing Sp6 viral RNA polymerase. The translational efficiency profile is similar to that derived from RNA transfection assay shown in C. (E) Effects of selected nNOS 5′-UTR sequences on translational efficiency assessed with a transient DNA transfection assay using SV40 promoter-directed luciferase reporter constructs. Luciferase activity was normalized to β-galactosidase activity and protein content. The translational efficiency profile is again similar to that derived from RNA transfection assay shown in C. Shown for B, C, D, and E are results from three or four identical experiments, each in duplicate (mean ± SE).
We also evaluated the effects of the various nNOS leader sequences on the translational efficiency of reporter constructs by using both RNA- and DNA-based transient transfections of nNOS(+) and nNOS(−) mammalian cells. C2C12 cells [mouse skeletal muscle, nNOS(+)], NT2/D1 cells [human neuroblastoma, nNOS(+)], and HeLa cells [human squamous carcinoma, nNOS(−)] were transiently transfected with capped chimeric nNOS 5′-UTR/luciferase reporter RNA and β-galactosidase RNA, both of which had been transcribed in vitro and quantified. Luciferase activity in cellular extracts, normalized for β-galactosidase activity and protein content, was determined on the linear portion of the activity/time after transfection curve. Again, nNOS leader sequences had important effects on the efficiency of translation (Fig. 3C). Several unique features were noted in the activity profiles. First, chimeric nNOS 5′-UTR/luciferase reporter RNAs were translated more efficiently in cells than in lysates. For instance, exon 2 nNOS sequences were translated 3- to 4-fold more efficiently in cells relative to the luciferase construct alone. Second, intriguing exon 1-specific translation effects were evident. As in lysates, leader sequences shown to be expressed in a cell-restricted fashion had contrasting effects on translation. Exon 1c repressed translation to a greater extent than exon 1a in both lysates and cells. This observation indicates that nNOS exon 1 variants that are expressed in the same tissue have contrasting effects on the efficiency of translation. Neuronal leader sequences (exons 1f and 1g, but not exon 1h) were more efficiently translated in cells as compared with lysates, yet exhibited leader-sequence-specific effects (compare exon 1f with 1g). When contrasted with luciferase alone, exon 1g was translated 30-fold more efficiently in cells versus lysates. It is intriguing that closely located exonic sequences that are under similar transcriptional control (exons 1f, 1g, and 1h are all enriched in neurons) differ markedly in terms of their effects on translation. Third, the profiles of translational efficiency of varied nNOS 5′-UTR sequences were similar in different cell types. This indicates the presence of cell-dependent, but not cell-specific, general factors governing translation.
Chimeric nNOS 5′-UTR/luciferase reporter expression was also examined by using liposome-mediated DNA transfections. Recombinant vaccinia virus expressing Sp6 viral RNA polymerase was used to infect cells transiently transfected with DNA plasmids encoding various exon 1/luciferase translational reporters. Findings were comparable to RNA-based cell transfections (Fig. 3D). We further evaluated the effects of DNA-based transfections by using the SV40 promoter/enhancer to direct nuclear transcription of chimeric nNOS 5′-UTR/luciferase. As shown in Fig. 3E, the translational efficiency of select exon 1 examples varied markedly in a variety of cell types. Taken together, these findings indicate the varied 5′-leader sequences have important effects on translational efficiency. The effects of nNOS leaders on translation were observed when the RNA was exogenously synthesized and presented (RNA transfections) or endogenously synthesized in either the cytoplasm (Sp6 polymerase-directed) or nucleus (SV40-directed).
Differences in the translational efficiency of the exon 1 variants may reflect sequence-specific ribonucleoprotein complexes. A 5′-UTR sequence-specific interaction with cellular RNA-binding proteins is therefore postulated for nNOS and can be modeled upon paradigms reported for ferritin and insulin-like growth factor II, among others (22–25). The current work did not seek to exhaustively characterize sites for transcription initiation for all nine exon 1 examples, but rather used representative cap sites characterized by 5′-RACE and/or RNase protection analysis. It may be anticipated that a further level of complexity will be superimposed by differences in transcription start sites across cell types expressing the same exon 1 or after cellular activation. Alternative sequences at the 5′ termini of any given exon 1 may include or exclude critical cis-RNA regulatory motifs. The existence of specific protein/RNA interactions modulating nNOS expression would provide a unique substrate for developmental pathways or allow differential responsiveness of nNOS leader sequences to changes in cellular phenotype. Consistent with this model, the translational efficiency of nNOS leader sequences differed markedly depending on cellular phenotype. The effects of the differentiation state of C2C12 skeletal muscle cells on the translational efficiency of exon 1a, 1b, and 1c chimeric nNOS 5′-UTR/luciferase reporter RNAs, all skeletal muscle-enriched exon 1 variants, were determined. The translation of 1a, but not 1b or 1c, was increased more than 100% in differentiated C2C12 cells (fused myotubes) compared with nondifferentiated C2C12 cells (myoblasts) (Fig. 4). No differences were observed in steady-state levels of reporter luciferase RNA as assessed by Northern blot analysis in this (Fig. 4) and prior transfections, indicating that these sequences did not affect the rate of degradation of the reporter RNA constructs. Therefore, the effect of nNOS leaders was at the level of translation of the chimeric RNAs. This may reflect the existence of a skeletal muscle cell-specific and differentiation-dependent translational cis-RNA element(s) within exon 1a.
Figure 4.

Effect of cellular phenotype on translational efficiency of selected nNOS 5′-UTR sequences. Translational efficiency of 1a was enhanced by more than 100% (P < 0.005) in differentiated vs. nondifferentiated C2C12 cells, whereas that of 1b and 1c was not affected. Shown are results from three identical experiments, each in duplicate (mean ± SE). Steady-state levels of luciferase reporter RNA were assessed with Northern blot analysis and was not affected by nNOS 5′-UTR sequences. Shown is a representative Northern result.
Discussion
This body of work emphasizes that temporal and spatial expression of the nNOS mRNA and protein is regulated at multiple levels: transcription, pre-mRNA splicing, and translation. Targeting of nNOS mRNA transcripts to diverse tissues, many of which are of great interest to the pathobiology of disease, is mediated by intriguing promoter diversity. Previous work had demonstrated that two of the exon 1 variants, namely 1f and 1g, are transcribed from different promoters (17, 18). We have found that nine unique exon 1 examples are utilized in establishing tissue-specific transcription of the human nNOS gene. The predominant molecular mechanism underlying these unique 5′ ends is alternative promoter usage. In each case the 5′ ends of nNOS mRNA species represent different first exons spliced to a common exon 2. This shared region encodes 3′ sequences of the 5′-UTR as well as the initiation codon with upstream stop codons in all three frames. Hence in this regard, one gene produces many different mRNA species that encode the same protein product.
Arguably, NOS1 is the most complex human gene yet described in terms of promoter diversity. The use of multiple promoters by a gene provides a genomic framework enabling a cell to respond to diverse physiological and pathophysiological stimuli and is a strategy utilized by a number of important genes, albeit in a simpler fashion. NOS1 exon 1 variants expressed in skeletal muscle or neuronal tissue were separately clustered in distinct genomic regions 75 kb apart. Such an unusual arrangement of promoters on the genome may indicate the existence of tissue-specific control mechanisms that serve to coordinate transcription initiation and elongation and perhaps foreshadows a sophisticated regulatory mechanism at the chromatin level. Exon 1-specific alternative splicing patterns were also observed for nNOS, implying that the pattern of splicing depends on the promoter structure from which the transcript originated, as described recently for fibronectin (26).
The alternative splicing patterns we have observed at the 5′ terminus of the human nNOS mRNA are complex and involve the deletion of exon 2 and/or the insertion of exon AS, both of which are associated with tissue and promoter specificity. Paradigms of alternative splicing patterns emphasize exonic or intronic RNA regulatory regions acting in either a positive (exon-selection) or negative (exon-skipping) fashion (27) coupled with hnRNPs (heterogeneous nuclear ribonucleoprotein particles) and tissue-specific RNA-binding proteins, such as NOVA (28) and Elav (29). Especially interesting in this respect is the unusually large size of exon 2 for an internal exon (1145 nt) in comparison with an average size of 137 nt in vertebrate genes (30). Exon 2 is flanked by very large introns, with intron 1 and 2 each ≥25 kb (17), and was the exon targeted by homologous recombination in a mouse model (31). Future studies seeking to define the mechanisms that control splicing patterns in the human nNOS will undoubtedly focus on this issue. The biologic consequences of these alternative splicing patterns were not experimentally addressed in the current study but are intriguing and merit further exploration. The deletion of exon 2 would remove the translation initiation codon for the full-length nNOS protein. Though it has been demonstrated that translation can initiate within exon 5, in both humans (32) and mice (3), the functional properties of the derived proteins (TnNOS/nNOSγ) remain to be exhaustively characterized. It should be noted that such nNOS variants would lack both the PDZ domain and the PIN binding motif important for distinct subcellular localization and in vivo enzymatic activity profiles, respectively (3, 33). The diversity of upstream RNA sequences observed in the current work indicates the potential existence of other nNOS protein variants. Notably, suitable in-frame upstream AUG initiation codons were found in exons 1a, 1b, and 1h if the transcripts lacked exon 2 (supplemental data, www.pnas.org). These variant nNOS proteins, if shown to exist, are related to the nNOSβ variant first noted in knock-out murine nNOSΔ/Δ models and may represent important physiological sources of NO in discrete brain regions (21).
Both alternative promoter usage and alternative splicing burden nNOS mRNA transcripts with complex 5′-UTRs. Such highly structured nNOS 5′ mRNA leader sequences present unique RNA sequences. Moreover, the human nNOS gene spans 240 kb of human genomic DNA and is therefore predicted to be kinetically inefficient in transcription initiation and elongation, requiring 1.6–2.5 hr, based on transcription velocity of 100–150 kb/hr (34). Therefore, the ability to recruit and promptly translate mature nNOS mRNAs may be more efficient than relying on nascent transcripts. Cell-dependent mechanisms appear dominant for in vivo translation of nNOS mRNA transcripts. For example, exon 1g is highly G+C rich and is predicted to exhibit extremely stable secondary structure (free energy −200.8 kcal/mol). Although exon 1g was translated extremely inefficiently in vitro, it was efficiently translated in all cell types tested. A sequence-specific interaction with an activating cellular factor(s) is postulated. Similar effects of a complex 5′-UTR have been reported for other genes: H-ferritin (22), c-myc (23), γ-glutamyltransferase (24), and insulin-like growth factor II (25). Thus, a complex 5′-UTR with many structural features is not merely a negative regulator of translation. Rather, it may be viewed as a substrate for different cellular factors to act upon in response to varied signaling pathways and may therefore represent a dynamic, central component of the translational control mechanism. We posit that these unique leader sequences contain cis RNA elements that modulate translational efficiency in a tissue- and differentiation-specific manner and therefore represent a novel regulatory mechanism in the NOS gene family. As an example, we have demonstrated that the translational efficiency of exon 1a is robustly enhanced upon myoblast fusion, whereas the translation of exons 1b or 1c remains unchanged. This effect is not mediated by changes in RNA stability. The role of cellular differentiation signals in selective modulation of translation efficiency of unique classes of RNA species is an exciting area for future study.
Newer facets of mRNA cellular biology, such as editing, frame-shifting, subcellular trafficking, and cis modifiers of translational efficiency and mRNA stability are exciting molecular determinants that may be involved in developmental cascades and specific disease processes. Given the protean roles ascribed for nNOS, it fits well that a multidimensional transcription and translation infrastructure is operative at the level of nNOS mRNA structure and function.
Supplementary Material
Acknowledgments
We thank Romesh Subramanian of Emory University for help with in situ hybridization. This work was supported by grants from the Heart and Stroke Foundation of Canada (T-3668, P.A.M.) and by the National Institutes of Health (HL48667, J.N.W.). Y.W. is a recipient of a Medical Research Council of Canada Fellowship Award. P.A.M. is a recipient of a Career Investigator Award from the Heart and Stroke Foundation of Canada.
Abbreviations
- nNOS
neuronal nitric oxide synthase
- UTR
untranslated region
- 5′-RACE
rapid amplification of 5′ cDNA ends
- RT-PCR
reverse transcription–PCR
- SV40
simian virus 40
Footnotes
References
- 1.Wang Y, Newton D C, Marsden P A. Crit Rev Neurobiol. 1999;13:21–43. doi: 10.1615/critrevneurobiol.v13.i1.20. [DOI] [PubMed] [Google Scholar]
- 2.Bredt D S, Hwang P M, Glatt C E, Lowenstein C, Reed R R, Snyder S H. Nature (London) 1991;351:714–718. doi: 10.1038/351714a0. [DOI] [PubMed] [Google Scholar]
- 3.Brenman J E, Chao D S, Gee S H, McGee A W, Craven S E, Santillano D R, Wu Z, Huang F, Xia H, Peters M F, et al. Cell. 1996;84:757–767. doi: 10.1016/s0092-8674(00)81053-3. [DOI] [PubMed] [Google Scholar]
- 4.Ahn A H, Kunkel L M. Nat Genet. 1993;3:283–291. doi: 10.1038/ng0493-283. [DOI] [PubMed] [Google Scholar]
- 5.Bentley D L, Groudine M. Mol Cell Biol. 1986;6:3481–3489. doi: 10.1128/mcb.6.10.3481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Eick D, Polack A, Kofler E, Lenoir G M, Rickinson A B, Bornkamm G W. Oncogene. 1990;5:1397–1402. [PubMed] [Google Scholar]
- 7.Vu T H, Martin G R, Lee P, Mark D, Wang A, Williams L T. Mol Cell Biol. 1989;9:4563–4567. doi: 10.1128/mcb.9.10.4563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Harada N, Utsumi T, Takagi Y. Proc Natl Acad Sci USA. 1993;90:11312–11316. doi: 10.1073/pnas.90.23.11312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nanbru C, Lafon I, Audigier S, Gensac M C, Vagner S, Huez G, Prats A C. J Biol Chem. 1997;272:32061–32066. doi: 10.1074/jbc.272.51.32061. [DOI] [PubMed] [Google Scholar]
- 10.Stein I, Itin A, Einat P, Skaliter R, Grossman Z, Keshet E. Mol Cell Biol. 1998;18:3112–3119. doi: 10.1128/mcb.18.6.3112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chen C Y, Del Gatto-Konczak F, Wu Z, Karin M. Science. 1998;280:1945–1949. doi: 10.1126/science.280.5371.1945. [DOI] [PubMed] [Google Scholar]
- 12.Wolf D A, Strobl L J, Pullner A, Eick D. Nucleic Acids Res. 1995;23:3373–3379. doi: 10.1093/nar/23.17.3373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kozak M. J Cell Biol. 1991;115:887–903. doi: 10.1083/jcb.115.4.887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sonenberg N. Gene Expr. 1993;3:317–323. [PMC free article] [PubMed] [Google Scholar]
- 15.Hentze M W, Kuhn L C. Proc Natl Acad Sci USA. 1996;93:8175–8182. doi: 10.1073/pnas.93.16.8175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Keilhoff G, Seidel B, Noack H, Tischmeyer W, Stanek D, Wolf G. Neuroscience. 1996;75:1193–1201. doi: 10.1016/0306-4522(96)00330-2. [DOI] [PubMed] [Google Scholar]
- 17.Hall A V, Antoniou H, Wang Y, Cheung A H, Arbus A M, Olson S L, Lu W C, Kau C L, Marsden P A. J Biol Chem. 1994;269:33082–33090. [PubMed] [Google Scholar]
- 18.Xie J, Roddy P, Rife T K, Murad F, Young A P. Proc Natl Acad Sci USA. 1995;92:1242–1246. doi: 10.1073/pnas.92.4.1242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wilcox J N, Subramanian R R, Sundell C L, Tracey W R, Pollock J S, Harrison D G, Marsden P A. Arterioscler Thromb Vasc Biol. 1997;17:2479–2488. doi: 10.1161/01.atv.17.11.2479. [DOI] [PubMed] [Google Scholar]
- 20.Lee M A, Cai L, Hubner N, Lee Y A, Lindpaintner K. J Clin Invest. 1997;100:1507–1512. doi: 10.1172/JCI119673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Eliasson M J, Blackshaw S, Schell M J, Snyder S H. Proc Natl Acad Sci USA. 1997;94:3396–3401. doi: 10.1073/pnas.94.7.3396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rogers J T, Andriotakis J L, Lacroix L, Durmowicz G P, Kasschau K D, Bridges K R. Nucleic Acids Res. 1994;22:2678–2686. doi: 10.1093/nar/22.13.2678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lazarus P. Oncogene. 1992;7:1037–1041. [PubMed] [Google Scholar]
- 24.Diederich M, Wellman M, Visvikis A, Puga A, Siest G. FEBS Lett. 1993;332:88–92. doi: 10.1016/0014-5793(93)80490-l. [DOI] [PubMed] [Google Scholar]
- 25.Nielsen F C, Ostergaard L, Nielsen J, Christiansen J. Nature (London) 1995;377:358–362. doi: 10.1038/377358a0. [DOI] [PubMed] [Google Scholar]
- 26.Cramer P, Pesce C G, Baralle F E, Kornblihtt A R. Proc Natl Acad Sci USA. 1997;94:11456–11460. doi: 10.1073/pnas.94.21.11456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Grabowski P J. Cell. 1998;92:709–712. doi: 10.1016/s0092-8674(00)81399-9. [DOI] [PubMed] [Google Scholar]
- 28.Buckanovich R J, Darnell R B. Mol Cell Biol. 1997;17:3194–3201. doi: 10.1128/mcb.17.6.3194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Good P J. Proc Natl Acad Sci USA. 1995;92:4557–4561. doi: 10.1073/pnas.92.10.4557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hawkins J D. Nucleic Acids Res. 1988;16:9893–9908. doi: 10.1093/nar/16.21.9893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Huang P L, Dawson T M, Bredt D S, Snyder S H, Fishman M C. Cell. 1993;75:1273–1286. doi: 10.1016/0092-8674(93)90615-w. [DOI] [PubMed] [Google Scholar]
- 32.Wang Y, Goligorsky M S, Lin M, Wilcox J N, Marsden P A. J Biol Chem. 1997;272:11392–11401. [PubMed] [Google Scholar]
- 33.Jaffrey S R, Snyder S H. Science. 1996;274:774–777. doi: 10.1126/science.274.5288.774. [DOI] [PubMed] [Google Scholar]
- 34.Tennyson C N, Klamut H J, Worton R G. Nat Genet. 1995;9:184–190. doi: 10.1038/ng0295-184. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}