

Abstract
Introduction:
An increasing number of patients present with multiple symptoms affecting many organs including the brain due to multiple mediators released by mast cells. These unique tissue immune cells are critical for allergic reactions triggered by immunoglobulin E (IgE), but are also stimulated (not activated) by immune, drug, environmental, food, infectious, and stress triggers, leading to secretion of multiple mediators often without histamine and tryptase. The presentation, diagnosis, and management of the spectrum of mast cell disorders are very confusing. As a result, specialists have recently excluded neuropsychiatric symptoms, and made the diagnostic criteria stricter, at the expense of excluding most patients.
Areas covered:
A literature search was performed on papers published between January 1990 and November 2018 using MEDLINE. Terms used were activation, antihistamines, atopy, autism, brain fog, heparin, KIT mutation, IgE, inflammation, IL-6, IL-31, IL-37, luteolin, mast cells, mastocytosis, mediators, myalgic encephalomyelitis/chronic fatigue syndrome, mycotoxins, release, secretion, tetramethoxyluteolin, and tryptase.
Expert opinion:
Conditions associated with elevated serum or urine levels of any mast cell mediator, in the absence of any comorbidity that could otherwise explain such increases, should be considered mast cell activation disorders, or better yet be collectively termed ‘Mast Cell Mediator Disorders (MCMD).’ Emphasis should be placed on the identification of unique mast cell mediators, and development of drugs or supplements that inhibit their release.
Keywords: Activation, antihistamines, autism spectrum disorder, brain fog, KIT mutation, cytokines, IgE, inflammation, IL-6, IL-37, luteolin, mast cells, mastocytosis, mediators, myalgic encephalomyelitis/chronic fatigue syndrome, mycotoxins, tetramethoxyluteolin, tryptase
1. Introduction
Over the last few years, there has been an unusual increase in atopic diseases, including allergies, asthma, eczema, rhinitis, and food sensitivities. In addition, there have been numerous cases of patients presenting with symptoms consistent with allergies, and/or inflammation, affecting many organs, but no recognizable trigger or too many non-allergic triggers. These symptoms (Table 1) originate from mediators secreted (released) from mast cells and include flushing, pruritus, hypotension, gastrointestinal complaints, headaches, irritability, malaise, memory loss, and neuropsychiatric issues. As a result, a number of different names have been proposed to capture these symptoms such as mast cell diseases or mast cell disorders (MCD) and mast cell activation diseases or mast cell activation disorders (MCAD).
Table 1.
Common Symptoms in Patients With Mast Cell Mediator Disorders
| • Cardiovascular: chest pain, hypotension, hypotensive syncope, tachycardia |
| • Dermatologic: angioedema, dermatographism, flushing, pruritus, urticaria pigmentosa |
| • Gastrointestinal: abdominal crumping/pain, bloating, diarrhea, esophagitis, nausea, vomiting |
| • Musculoskeletal: bone/muscle pain, degenerative disc disease, osteoporosis/osteopenia |
| • Naso-ocular: nasal congestion, pruritus, tearing |
| • Neurologic: headache, memory and concentration difficulties (brain fog), paresthesias, peripheral neuropathy |
| • Respiratory: hoarseness, sore throat, stridor, throat swelling, wheezing |
| • Systemic: anaphylaxis, fatigue |
Unfortunately, these are many confusing aspects both in the naming and diagnosis of such disorders. First, the term ‘activation’ is typically reserved for activation of receptors or enzymes while ‘stimulation’ is used for cells. Second, stimulation of mast cells may imply proliferation without secretion of all or even any mast cell mediators as in the case of mast cell sarcoma. Next, the term ‘secretion’ is usually reserved for mediators stored inside secretory granules, while the term ‘release’ is used for both pre-stored and newly synthesized mediators. Be it as it may, release of mast cell mediators can occur in many physiologic and pathologic settings. To make matters worse, there are a number of other diseases that mimic and/or are comorbid with diseases involving mast cells (Table 2). As a result, affected patients often go for 10–20 years and as many physicians of different specialties before a diagnosis is made, by which time they have been prescribed numerous medications, often with severe drug interactions that further complicate their presentation and course of their disease.
Table 2.
Conditions Often Comorbid With Mast Cell Diseases
| • Chronic inflammatory response syndrome (CIRS) |
| • Fibromyalgia syndrome (FMS) |
| • Ehlers-Danlos Syndrome (EDS) |
| • Gulf War Illness (GWI) |
| • Interstitial cystitis/bladder pain syndrome (IC/BPS) |
| • Irritable bowel syndrome (IBS) |
| • Kounis syndrome |
| • Multiple chemical sensitivity syndrome (MCSS) |
| • Myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) |
| • Post-Lyme syndrome |
| • Postural orthostatic tachycardia syndrome (POTS) |
| • Post-traumatic stress disorder (PTSD) |
2. Mast cell biology
Mast cells are immune cells derived from hematopoietic precursors and mature in tissue microenvironments [1]. Mast cells are critical for the development of allergic reactions [2], but also act as sensors of environmental and psychological stress [3,4]. Perivascular mast cells were shown to probe cutaneous blood vessels by extending philopodia through endothelial gaps and capture immunoglobulin E (IgE) from blood vessels [4], a process that might also allow mast cells to sense and capture other circulating molecules. Mast cells also participate in the innate immune system (host defense against infectious pathogens, neutralization of toxins) and in the adaptive immune response [5–9], but also in antigen presentation [10,11] regulation of T cell responses [12–14], even in the absence of antigen presentation [15], autoimmunity [16], and inflammation [17–19].
Mast cells are located in tissues that interface with the external environment [20] and are stimulated primarily by IgE-dependent reactions (allergen). In addition to allergens, mast cells are also stimulated by a variety of different triggers (Table 3) that include bacteria, drugs, foods, fungi, heavy metals, organophosphates, viruses, and ‘danger signals’ [2], as well as certain neuropeptides including corticotropin-releasing hormone (CRH) [21], neurotensin (NT) [22,23], and substance P (SP). Both NT [24,25] and SP [26–29] are known to participate in inflammatory processes. Many cationic drugs have now been shown to stimulate mast cells through activation of the low affinity G-protein-coupled receptor MRGPRX2 [30]. Mast cells commonly respond to non-allergic triggers leading to mediators may also be released selectively without degranulation making it difficult to identify these mast cells with routine histology [31] (Table 4).
Table 3.
Cell Triggers of Mast Cell Degranulation
| Acetylcholine |
| Complement fragments |
| - C3α, C4α, C5α |
| Drugs |
| - Local anesthetics |
| - Lactam antibiotics |
| - Neuromuscular junction blockers |
| - Vancomycin |
| IgE |
| IgG1 |
| IgG4 |
| Lysophosphatidylserine |
| Peptides |
| - Adrenomedullin |
| - CGRP |
| - Endorphin |
| - Endothelin |
| - Eosinophil granule proteins |
| - Hemokinin-1 |
| - Leptin |
| - Mastoparan |
| - Neurotensin |
| - NGF |
| - PTH |
| - Somatostatin |
| - SP |
| - Thrombin |
| - VIP |
| Physical conditions |
| - Cold |
| - Heat |
| - Pressure |
| - Stress |
| - Vibration |
Table 4.
Triggers of Mast Cells Without Degranulation
| Triggers | Mediator |
|---|---|
| Peptides | |
| CRH | VEGF |
| SCF | IL-6 |
| Cytokines | |
| IL-1β | IL-6 |
| IL-33 | IL-31 |
| IL-33 | CX CL8 (IL-8) |
| IL-33 + SP | TNF, VEGF |
| Heavy metals | |
| Aluminum | |
| Cadmium | |
| Mercury | |
| Herbicides | |
| Atrazine | |
| Glyphosate | |
| Pathogens | |
| Borrelia (Lyme disease)* | TNF |
| LPS | TNF |
| Poly (I:C) (viruses) | IL-6, TNF |
| Sporothrix (mold)* | IL-6, TNF |
Mycotoxins
Upon stimulation, mast cells secrete preformed mediators (Table 5) such as β-hexosaminidase (β-hex), histamine, tumor necrosis factor (TNF), and tryptase through rapid (1–5 min) degranulation. Mast cells are the only cell type that stores preformed TNF [32], which is rapidly secreted and influences T cell recruitment and activation [33–35]. Mast cells also release newly synthesized phospholipid products such as prostaglandin D2 (PGD2) and leukotrienes [36–38]. Mast cells also secrete numerous de novo synthesized protein mediators typically 6–24 h after stimulation such as cytokines [39], chemokines (TNF, CCL2, CCL8), and peptides hemokinin-1 (HK-1), and renin [19,40] (Table 4). Mast cell-derived CCL2 and CXCL8 enhance recruitment of other immune cells to the site of inflammation [41].
Table 5.
Mast Cell Mediators
| Prestored |
| Biogenic Amines |
| Dopamine |
| Histamine |
| 5-Hydroxytryptamine (5-HT, serotonin) |
| Polyamines |
| Spermidine, spermine |
| Cytokines |
| TNF |
| Enzymes |
| Arylsulfatases A |
| Beta-hexosaminidase |
| Beta-glucuronidase |
| Beta-glucosaminidase |
| Beta-D-galactosidase |
| Carboxypeptidase A |
| Cathepsins B,C, D, E, L |
| Chymase |
| Garnzyme B |
| Kinogenases |
| Phospholipases |
| Renin |
| Tryptase |
| Metalloproteinases |
| (CPA3, MMP9, ADAMTSS) |
| Growth factors |
| FGF |
| NGF |
| SC |
| TGFβ |
| VEGF |
| Peptides |
| ACTH |
| Angiogenin |
| Angiopoietin |
| Calcitonin gene-related peptide |
| Corticotropin-releasing hormone |
| Endorphins |
| Endothelin |
| Hemokinin-1 |
| Kinins (bradykinin) |
| Leptin |
| Melatonin |
| Neurotensin |
| RANKL |
| Somatostatin |
| Substance P |
| Urocortin |
| Vasoactive intestinal peptide |
| Proteoglycans |
| Chondroitin sulfate |
| Heparan sulfate |
| Heparin |
| Hyaluronic acid |
| Serglin |
| De novo synthesized |
| Chemokines |
| IL-8 (CXCL8), MCP-1 (CCL2), MCP-3 (CCL7), |
| MCP-4, RANTES (CCL5), Eotaxin (CCL11) |
| Cytokines |
| IL-1β, IL-4, IL-5, IL-6, IL-15, IL-17, IL-31, IL-33, TNF |
| Growth Factors |
| SCF, β-FGF, neurotrophin 3, NGF, |
| PDGF, TGFβ, VEGF |
| Nitric oxide |
| Phospholipid metabolites |
| Leukotriene B4 |
| Leukotriene C4 |
| Platelet activating factor |
| Prostaglandin D2 |
Moreover, corticotropin-releasing hormone (CRH or factor, CRF) is also synthesized by mast cells [42] implying that it could have autocrine effects [17,43]. In particular, CRHR-1 is expressed on human cultured mast cells, activation of which induces production of vascular endothelial growth factor (VEGF) without tryptase [21].
Mast cells can secrete IL-31, which is particularly pruritogenic [44], as well as additional ‘danger’ signals [45] (Table 4). Mitochondrial DNA (mtDNA) [34,46] can lead to auto-inflammatory responses [47–50], augment allergic responses [51], and has direct neurotoxic effects [52]. We had reported that mtDNA is increased in the serum of children with autism spectrum disorder (ASD) [53]. Another key danger signal is the alarmin IL-33 [54], which is secreted by fibroblasts and endothelial cells, and has been implicated in many allergic [55] and inflammatory [56] diseases. IL-33 augments the effect of IgE on secretion of histamine from mast cells and basophils [54,57]. It is interesting that the most widely used herbicide glyphosate induces IL-33 expression and airway inflammation [58].
We showed that IL-33 augments the ability of SP to stimulate secretion of VEGF from human mast cells without degranulation [59]. We recently also reported that SP and IL-33, when administered in combination, lead to an impressive increase in the gene expression and secretion of TNF [60] and IL-1β [61] from cultured human mast cells. Mast cells can secrete IL-33 [62,63], as well as the SP-related peptide HK-1 [64], implying autocrine augmentation. Moreover, tryptase secreted from mast cells acts on extracellular IL-33 and generates mature, more active IL-33 [65], which then stimulates mast cells to secrete IL-1β, which in turn stimulates mast cells to secrete IL-6 [66]. In addition, mast cell-derived TGFβ promotes the development of Th17 cells and mast cells can also secrete IL-17 themselves [67].
Stimulated mast cells can secrete their numerous bioactive mediators [19,68,69] utilizing different signaling [70–73] and secretory [70,74] pathways. One important pathway is that of mammalian target of rapamycin (mTOR) [75], which we have shown to be stimulated by SP in human cultured mast cells [76]. The ability to secrete multiple mediators allows mast cells to actively interact with other cell types in their surrounding environment, especially T cells [77,78]. It is interesting that secretion of mast cell mediators have been shown to be regulated by a circadian clock [79,80].
Each mediator could lead to specific clinical features. For instance, histamine is associated with headaches, hypotension, and pruritus; tryptase with inflammation and fibrinogen lysis; cytokines and chemokines with constitutional symptoms of generalized inflammation and fatigue, PGD2 with flushing, and leukotrienes with bronchoconstriction. As a result, mast cell-derived mediators could contribute to the pathogenesis of not only allergic diseases, asthma and mastocytosis [2], but also in atopic dermatitis, psoriasis, myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) [81], fibromyalgia syndrome (FMS) [82], coronary artery disease, and obesity [83], as well as ASD [17,50,84].
The severity of symptoms depends on many factors, such as the capacity of mast cells to secrete mediators, the levels of circulating IgE, the presence of cytokines and chemokines [85], especially IL-33 [59], as well as the co-existence of high-risk conditions, such as stimulation by hymenoptera venom(s) especially in patients suffering from mastocytosis [86–89]. Reactivity to wasp stings could lead to anaphylaxis, characterized by the sudden onset of severe flushing, swelling of the throat, bronchoconstriction, and hypotension that may progress to death [90]. Basophils also participate in allergic and other inflammatory reactions in-ways somewhat similar to that of mast cells, except that their main trigger is IgE and they do not contain tryptase [91,92]. Various assays have been proposed to document the involvement of basophil activation [93,94], via identification of CD63 and CD203c [93–95].
3. Classification and diagnostic features of mast cell disorders
This topic is quite confusing because of the use of different terminology by various specialists and consensus groups. For instance, the terms ‘mast cell diseases,’ ‘mast cell disorders,’ and ‘mast cell syndromes’ are often used interchangeably. In this review, the term ‘mast cell disorders’ is used as it includes many variants or subtypes.
Disorders involving mast cells [96,97] have been traditionally classified into three general categories: primary, secondary, and idiopathic [98,99]. However, new information on the genetic and epigenetic causes of the occurrence and severity of the symptoms [100] requires a new approach to the understanding and classification of these disorders as shown in Table 6. Historically, ‘primary’ has been used to imply that these disorders are caused by genetic and/or epigenetic alterations leading to functional consequences in proliferation of mast cells in the bone marrow, skin, or other tissues/organs.
Table 6.
Classification of Mast Cell Disorders
| Clonal Mast Cell Proliferation Disorders |
| 1. Systemic mastocytosis (indolent, aggressive) |
| 2. Cutaneous mastocytosis (urticarial pigmentosa, diffuse, telangiectasia macularis eruptive persistans) |
| 3. Mast cell leukemia (MCL) |
| 4. Mast cell sarcoma (MCS) |
| 5. Extracutaneous mastocytoma (benign) |
| 6. Monoclonal mast cell activation syndrome (MMAS) |
| Mast Cell Mediator Disorders (Idiopathic) |
| 1. Monoclonal without abnormal clustering |
| 2. Non-clonal |
| 3. Idiopathic anaphylaxis (IA) |
| Reactive Mast Cell Proliferation/Mediator Release Disorders |
| 1. IgE-mediated hypersensitivity reactions (e.g. food, insect anaphylaxis) |
| 2. Drug-induced (e.g. vancomycin, opioids, taxanes, muscle relaxants, adenosine, nonsteroidal anti-inflammatory) |
| 3. Mast cell hyperplasia (related with chronic infections, neoplasia, autoimmune conditions due to a possible excess of stem cell factor) |
3.1. Clonal mast cell proliferation disorders
3.1.1. Systemic mastocytosis
Systemic mastocytosis (SM) is defined by mast cell involvement of at least one extracutaneous tissue typically in the bone marrow, if certain criteria are fulfilled [101]: one major criterion of mast cell clusters in bone marrow biopsy and one minor criterion (e.g. elevated basal serum tryptase, spindle-shaped mast cells), or three minor criteria established by the World Health Organization (Table 7) [102]. However, these criteria are strongly based on the presence of a ‘gain-of-function’ mutation in the gene KIT at codon 816 (D816V) or other codons nearby. Systemic mastocytosis can be characterized as (a) indolent, (b) aggressive, or (c) associated with a hematologic non-mast cell lineage disease. Sometimes a single bone marrow biopsy may miss mast cell clusters. In our study, only 17% of patients had evidence in one biopsy, but 83% of patients had evidence of mastocytosis when two iliac crest biopsies are taken [103].
Table 7.
Diagnostic Criteria for Systemic Mastocytosis
| Major criterion | a. Multifocal, dense infiltrates of mast cells (≥15 mast cells in aggregates) detected in intramedullary biopsy sections and/or extramedullary organ(s). |
| Minor criteria | a. In intramedullary biopsy sections or other extramedullary ones >25% of the mast cells in the infiltrate are spindle shaped or have atypical morphology, or of all mast cells in bone marrow aspirate smears >25% are immature or atypical |
| b. Gain of function point mutation of KIT at codon 816 (usually KIT D816V) in bone marrow, blood, or other extracutaneous organ | |
| c. Aberrant immunophenotype of mast cells of CD2 and/or CD25 in bone marrow, blood, or other extracutaneous organ (in addition to normal mast cell markers) | |
| d. Persistently elevated baseline serum total tryptase (>20 ng/ml). |
Mast cells should be identified by immunohistochemistry for both tryptase and KIT (CD117), the surface receptor for stem cell factor (SCF), because KIT indicates the presence of all mast cells whether they have degranulated or not while the presence of tryptase or lack thereof indicates the degree of degranulation. CD117 is considered the marker of choice for the GI tract, especially in the portal triads. Flow cytometric immunophenotyping using antibodies against tryptase, CD117 (KIT), and CD25 could be performed in peripheral blood in suspected cases before advancing to bone marrow biopsies (Figure 1). Mast cells are low at bone marrow smears, and many cells are damaged making it difficult to identify abnormal mast cells.
Figure 1.
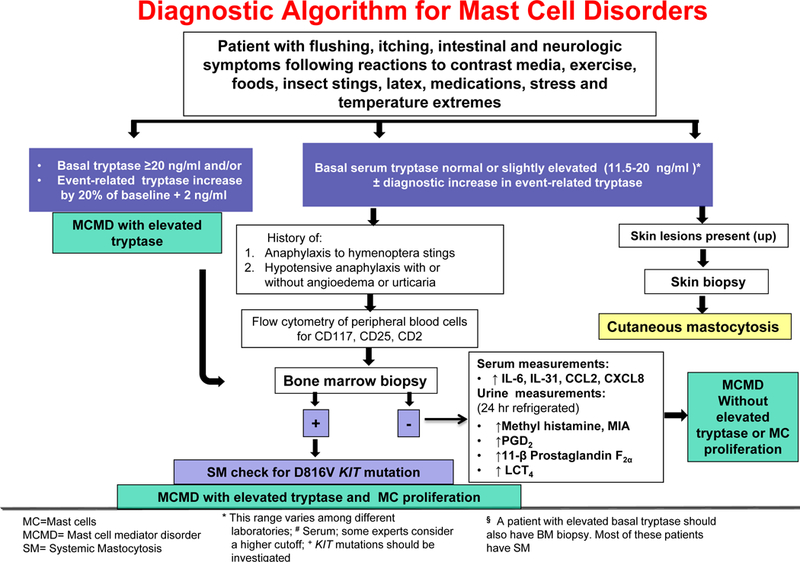
Diagrammatic representation of a proposed diagnostic rubric for mast cell mediator disorders with emphasis on clonal and non-clonal subtypes, and the presence or absence of elevated serum tryptase.
Typically, patients are considered candidates for bone marrow biopsy is: (a) Tryptase >20 ng/ml; (b) have a history of previous hypotensive episodes and/or syncope; (c) diffuse cutaneous mastocytosis (CM) in adult patients; (d) unexplained anaphylaxis or explained anaphylaxis provoked with hymenoptera stings even when specific IgE testing to hymenoptera is negative [104–106]. Bone marrow or peripheral mast cells should then be tested for the presence of the KIT mutation D816V, which is present in over 90% of patients with SM [107]: (e) Insect-induced anaphylaxis with symptoms different than those of SH [108]; (f) Presence of unexplained osteoporosis, hepatomegaly, and splenomegaly (suspicion for an aggressive variant of SM) [104]; (g) Abnormalities on peripheral blood count in patients with a MCAD (several hematologic non-mast cell lineage such as chronic myelomonocytic leukemia and myelodysplastic syndrome, but also other myeloproliferative neoplasms and lymphoproliferative diseases) [109] (Figure 1).
The most common mutation found in patients with SM is the point mutation located at codon 816 of KIT which affects mast cells: (a) It promotes the autonomous growth and expansion of the neoplastic mast cells [110]; (b) It can trigger mast cells to release IL-6 at high concentrations of SCF [111]. The D816V mutation may also be expressed in individuals in whom criteria for SM are not fulfilled [86,99,106,112–114], in which cases, the patients are considered to have monoclonal mast cell activation syndrome (MMAS) (see below). There are some additional, but not as common, functionally activating mutations of KIT (e.g. D419H, V560G), which do not induce detectable morphological alterations of affected mast cells; these mutations can still influence the evolution of the disease and/or the manifestation of certain symptoms [59].
Some other gene mutations (e.g. FIP1L1-PDGFRA) result in a constitutively activated platelet-derived growth factor receptor-α (PDGFRA) and have been associated with a primary eosinophilic disorder. In one study, 10 (56%) of 19 patients with systemic mast cell disease associated with eosinophilia carried this specific mutation, which when present is indicative of underlying systemic mastocytosis [115].
3.1.2. Cutaneous mastocytosis
In this subcategory, affected skin most commonly presents as urticaria pigmentosa (UP) or diffuse CM, and less frequently as bullous CM or solitary mastocytoma [96,116]. Different from adults with CM, bone marrow biopsy is not recommended for children [117,118]. It is generally considered that pediatric CM regresses by puberty in most children [116], but there have not been any systematic longitudinal studies to prove this point. However, increased serum baseline total tryptase identifies children at risk for SM [119], as does the presence of maculopapular cutaneous mastocytosis [120] [121] or detection of KIT D816V in peripheral blood [122]. It is interesting that KIT D816V is not present in most skin lesions suggesting that there may be committed stem cells outside the bone marrow. In this case, the molecules triggering mast cell proliferation in the skin may be NGF [123], RANTES [124], or even CRH acting on precursors in the hair follicle mesenchyme [125]. It is interesting that new UP lesions in children often occur in areas of trauma.
Telangiectasia macularis eruptive persistans is a variant of CM, not associated with urticaria [126].
3.1.3. Monoclonal mast cell activation disorder
This subcategory is limited to patients with systemic reactions to hymenoptera stings, who rarely have increased baseline serum tryptase, or in patients with unexplained episodes of anaphylaxis [127]. In these patients, bone marrow biopsy detects monoclonal mast cells that carry the mutant KIT D816V also detectable in bone marrow aspirate [128], and/or aberrant expression of CD2 or CD25. However, criteria for diagnosis of SM are not fulfilled, as only one or two minor criteria are met [104].
In conclusion, while symptoms due to release of mast cell mediators are present in most of the primary mast cell disorders, they also differ considerably: (a) Multifocal bone marrow mast cell aggregates and possible UP are only found in patients with SM; (b) D816V KIT mutation and aberrant CD25 expression on bone marrow mast cells characterizes patients with SM and monoclonal mast cell activation disorder (MMAD); (c) elevated baseline tryptase, and elevated baseline 24-h urine methylhistamine or PGD2/11β-PGF2α is found in most affected patients; (d) The response to anti-mediator therapy is variable and likely ineffective in patients with idiopathic anaphylaxis (IA) [99].
3.2. Reactive mast cell disorders
In these patients, no mutation of KIT D816V is found and flow cytometry is often negative for the expression of CD25 [98].
3.2.1. IgE-mediated hypersensitivity reactions
These are typical reactions (e.g. food, insect), due to aggregation of high-affinity IgE receptors (FcεRI) by allergen-bound IgE as was discussed earlier.
3.2.2. Drug-induced
Mast cells can respond to numerous drugs such as muscle relaxants, nonsteroidal anti-inflammatory, opioids, taxanes, and vancomycin. Such drugs can stimulate mast cells either directly or indirectly through G-coupled receptors [129]. It was recently discovered that the MRGPRX2 receptor is responsible for pseudo-anaphylactic reactions to many cationic drugs [30].
3.2.3. Mast cell hyperplasia
Hyperplasia in this context may be defined as increased disseminated mast cell density. This subvariant includes different diseases such as chronic infections, neoplasia, and autoimmune conditions that can stimulate mast cells through different mechanisms [98]. For instance, psoriasis is characterized by aberrant IFN-γ, which triggers mast cells through upregulation of high-affinity IgG receptors [130]. Mast cells in patients with rheumatoid arthritis are stimulated via C3a and C5a by binding to their respective receptors [130]. In patients with infectious diseases, mast cells are stimulated through the direct activation of Toll-like receptors (TLR) [131] that recognize molecular patterns common to microbial or viral pathogens.
However, there have been a number of reports of increased number of mast cells in the intestine of patients with SM [132], but also in the absence of SM in which case they may be termed ‘intestinal mastocytosis’ [133].
4. Mast cell mediator disorders
4.1. Clonal mast cell proliferation disorders
Most of these patients could have symptoms consistent with release of mast cell mediators.
4.2. Non-clonal mast cell mediator disorders
Most patients fall under this category that has also been called non-clonal mast cell activation disorder (nc-MCAS), mast cell activation syndrome (MCAS), or idiopathic MCAS[127] (Figure 2) depending on whether specific criteria are fulfilled [99,134–136]:
Figure 2.
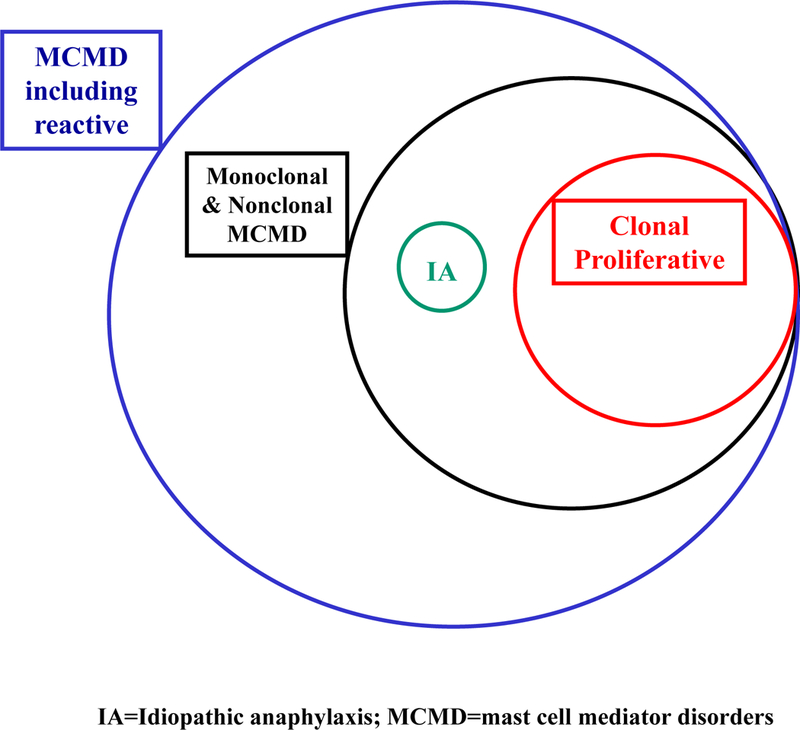
Estimated occurrence and overlapping nature of mast cell mediator disorders with respect to clonal and non-clonal subtypes: (a) clinical signs and symptoms; (b) documentation of increased serum and/or 24 h urine mast cell mediators or their metabolites; and (c) reduction in symptoms following treatment with drugs that inhibit stimulation of mast cells or the action of their mediators.
The second criterion listed above was recently modified [137,138] by excluding neurological symptoms, and emphasizing serum tryptase levels invariably leading to two subclasses of such patients, with or without serum tryptase elevations. In contrast, the symptom surveys of the mastocytosis society noted that >90% of patients reported ‘brain fog’ [139] as their major complaint and ‘stress’ as a key trigger [140,141]. Neuropsychiatric symptoms have been reported on many occasions [142–145]. Moreover, patients with mast cell disorders are profoundly affected by emotional stress via CRH [21,33,38,43]. In one case, UP expanded dramatically after emotional stress [146], while in another bone marrow mast cells were shown to express CRHR-1 [147]. In addition to the potential triggers already discussed, one should include the effects of mold and mycotoxins in neuropsychiatric diseases [148,149]. For instance, yeast stimulates mast cells to secrete IL-6 and TNF without degranulation [150] and so does Borrelia burgdoferi spirochetes [151].
The disorders in this subcategory have no identifiable causes, no allergen-specific IgE and no detectable clonal mast cells, based on D816V KIT mutation. Environmental or endogenous stimuli, intrinsic mast cell defects may be involved. The best way to describe the status of the mast cells is that they are ‘unstable,’ exhibiting aberrant stimulation [152]. This ‘unstable’ status of mast cells may be affected by positive and negative signals [153], lower stimulation threshold, or even ‘normal’ secretion of mast cell mediators, but with an abnormal response of the surrounding tissues (e.g. deficiency of catabolic enzymes such as diamine oxidase and histamine N-methyltransferase). Moreover, unstable mast cells may retain a ‘metabolic memory’ of past triggers that prime mast cells to respond more rapidly and more severely to exposure of the same or different triggers even though the original trigger may no longer be present. One such example would be the ability of stress via CRH to prime mast cells for the action of IgE [51] or NT [23].
Diseases such as carcinoid syndrome, pheochromocytoma, gastrinoma, and VIPoma, which could explain many of the symptoms have to be ruled out through specific diagnostic tests. For instance, serum chromogranin A is a biomarker for carcinoid syndrome, but not for SM [154]. If mast cell clonality is absent, mast cell mediators (e.g. serum tryptase, and 24-h urine N-methylhistamine, PGD2, 11β-PGF2α ) are measured in symptomatic patients (Figure 1). If these are not elevated, further clinical evaluation is necessary [118]. Idiopathic MCAS may be an underlying cause of various other frequent clinical presentations, like in subsets of patients with ME/CFS [81], fibromyalgia syndrome [82], and IBS [155,156].
‘Activation’ is usually inferred by release of mast cell mediators [157], primarily focusing on increased serum tryptase levels. However, serum tryptase levels could be both inadequate and misleading as mast cells can secrete many mediators especially cytokines and chemokines without any tryptase [31,60,61]. Tryptase is a key specific mediator for mast cells and it is recommended as the marker of choice, even though it is also released in small quantities by basophils [158,159]. Serum tryptase is usually detected with the B12 monoclonal antibody (mAb), which binds both alpha and beta tryptase. The G5mAb is bound with 10× more affinity to beta tryptase, so the alpha one is calculated as the difference between total and beta tryptase. The minimal increase in serum tryptase required as indicative of mast cell activation has been reported to be defined from the following equation: 20% of baseline tryptase level + 2 ng/ml + the baseline tryptase level = the value above which the definition of mast cell activation is met [98], usually ≥ 20 ng/ml. However, the sensitivity of this algorithm decreases with increased interval between resolution of symptoms and blood-draw [138]; moreover, there is no published data supporting this equation as has been extensively discussed in the past [160].
In addition, tryptase measurement could be affected by several conditions leading to false positive results: (a) presence of rheumatoid factor, (b) measurement of serum tryptase within 0.2–4 h after appearance of symptoms [98] if a symptomatic baseline level of serum tryptase is available, then a rapid increase is considered a reliable index; (c) the baseline has to be assessed after the patient’s complete recovery or during a symptom-free interval at least 24–48 h after complete resolution of all symptoms [161–163]; and (d) the sensitivity of tryptase algorithm decreases with decreasing clinical severity [138].
Histamine is specific for mast cells, but serum levels are not useful because histamine is metabolized very quickly (<1 min). Moreover, serum levels may be increased due to (a) release from basophils [158,159]; (b) tissue injury; (c) venous stasis, hemolysis or clotting during blood collection and sample preparation, or improper centrifugation/storage; (d) a diet rich in histamine [164]. The most informative test is measurement of the metabolite N-methylhistamine (30% of histamine) or 1-methyl imidazole acetic acid, MIA (60% of histamine) in 24 h urine or first morning urine collection, but must be kept cold during collection and sent to the laboratory cold; moreover, the patient should avoid consumption of foods containing biogenic amines (e.g. bananas, cheeses, spinach, sour crow, cumin, and other spices for 24 h before blood draw).
Serum PGD2 may be useful, but its release can come from (a) cellular elements through processing of peripheral blood samples; (b) due to a diet rich in niacin or niacin products used to treat atherosclerosis [165]; and (c) there are no established criteria for its normal serum levels. Moreover, PGD2 is rapidly metabolized in blood and it should best be detected by measuring its metabolite 11β-PGF2αas was discussed earlier. One paper reported that out of 25 patients studied with MCAS, 24 urine 11β-PGF2α was elevated in most and best correlated with flushing and itching [166].
Heparin is a specific mast cell mediator, and its increase in serum of SM patients has been documented [167].
In the absence of any inflammatory disease, serum IL-6 has been shown to correlate with disease severity and prognosis in SM [168–170]. A recent paper reported that increased serum levels of CCL2 strongly correlated with aggressive SM [171]. We recently reported impressive secretion of IL-31 from cultured human mast cells stimulated by IL-33 with or without SP or IgE [44]. Increased levels of SP were reported in the serum of SM patients [172].
4.3. Idiopathic anaphylaxis
These patients are characterized by (a) life-threatening anaphylactic reactions; (b) increased baseline levels of tryptase between the episodes; (c) absent mast cell clonality; and (d) acute onset of the disease. Anaphylaxis may be different in patients with IA (urticaria is present and serum IgE is significantly elevated) and patients with indolent SM (urticaria is absent during anaphylactic episodes and serum tryptase levels are significantly elevated) [112]. Recent evidence indicates that dendritic cells may deliver allergens to mast cells in the skin via extracellular microvesicles (exosomes) [173]. Evidently, mast cells can also secrete extracellular microvesicles [174] that may be directed specifically to T cells [175].
5. Histamine sensitivity
One must also consider patients who are either sensitive to histamine or have mutations in the gene diamine oxidase rendering them unable to metabolize histamine.
6. Activation threshold
There can be many reasons why mast cells can be stimulated to release their mediators (Table 8). Combination of triggers is more important than individual ones as they can lower the stimulation threshold of mast cells and ‘prime’ them for additional triggers [176] (Table 8). There are numerous examples of synergistic effects of different triggers. We reported that combination of CRH and NT have synergistic action in stimulating VEGF secretion without tryptase [23], as well as induce the expression of each other’s receptors on human cultured mast cells [177]. We also showed that NT [177] and SP [178] increase expression of CRHR-1 on human mast cells. Moreover, SP increases expression of the IL-33 receptor ST2 and IL-33 increases NK-1 receptor on human mast cells [60]. Stimulation of mast cells by SCF also cross-activates ST2 [179].
Table 8.
Proposed Pathogenetic Mechanisms for Mast Cell Mediator Disorders
| Lower activation threshold due to decreased expression/function of inhibitory molecules/pathways. |
| Intracellular |
| PTEN ↓ |
| Chondroitin sulfate ↓ |
| Sergylin ↓ |
| Spermine/spermidine ↓ |
| TGF beta ↓ |
| Extracellular |
| Deflisinc ↓ |
| IL-10 ↓ |
| IL-37 ↓ |
| IL-38 ↓ |
| TGF beta ↓ |
| Combination of non-allergic triggers |
| Peptides and Cytokines |
| SP + IL-33 |
| NT + CRH |
| HK-1 + IL-33 |
| Unappreciated triggers |
| CRH |
| Borrelia toxin |
| Bradykinin |
| CGRP |
| RCAP |
| Mycotoxins |
| NGF |
| Poly (lic) |
| PTH |
| Stress |
| Autocrine effects |
| Secretion of autocrine mediators |
| CRH |
| NK-1 |
| Neurotensin |
| Tryptase |
| Autocrine receptor expression |
| Secretion of autocrine mediators |
| NT → CRHR-1 ↑ |
| SP → CRHR-1 ↑ |
| CRH → NTR ↑ |
| SP → STZ ↑ |
| IL-33 → NK-1 ↑ |
7. Comorbidities
Symptoms alone and response or lack thereof to medication is not sufficient for diagnosis as many of the symptoms could derive from other pathologic entities, whether they implicate mast cells or not. Nevertheless, the presence of multisystemic manifestations of symptoms at the same time and in the absence of any other systemic disease is highly suspicious for the presence of some mast cell disorder. Symptoms suggestive of mast cell disorders that could be due to other conditions include: (a) ‘flushing’ is associated also with menopause, carcinoid syndrome, pheochromocytoma, medullary carcinoma of the thyroid gland, as well as ingestion of niacin; (b) cardiovascular disturbances may be related to myocardial infarction, endocarditis, autonomic dysfunction (dysautonomia), and postural orthostatic tachycardia syndrome (POTS), (c) gastrointestinal problems may implicate irritable bowel syndrome (IBS), food toxicity especially from spoiled scombroid fish (e.g. tuna) that generates large amounts of histamine, or neuroendocrine tumors such as VIPoma; (d) neurologic symptoms may be associated with panic attacks, migraines, epilepsy, and central nervous system (CNS) tumors; (e) respiratory signs may be part of asthma, ‘heartburn,’ angioedema; and (f) skin symptoms may be associated with acute toxic dermatoses, psoriasis, high parathyroid hormone (PTH), pemphigus vulgaris, and systemic lupus erythematosus (SLE). Other comorbid diseases include coronary hypersensitivity [180,181], and multiple chemical sensitivity syndrome (Table 1) [182]. Two particularly relevant diseases are Myalgic Encephalomyelitits/Chronic Fatigue Syndrome (ME/CFS) and ASD [81], both of which have recently been associated with focal inflammation of the brain [183].
We had reported that children with mastocytosis have a 10-fold higher risk of developing ASD [184]. ASD is a pervasive neurodevelopmental disorder characterized by deficits in communication, as well as the presence of restricted, repetitive behaviors [185–187]. Allergic symptoms, due to mediators secreted from mast cells, have been significantly correlated with ASD severity [183,188]. A recent paper reported that diffuse cutaneous mastocytosis with the novel somatic KIT mutation K509I was associated with tuberous sclerosis, most of the may affected patient of which have ASD [189]. Large epidemiological studies have reported that allergies and asthma in preschoolers are significantly associated with ASD [190–193]. A similar conclusion was reached in a systematic review showing a significant association between atopic dermatitis and ASD [194]. Moreover, the presence of allergic symptoms strongly correlated with the presence of serum autoantibodies against brain peptides [195,196] in children with ASD [197].
Inflammation is a complicated immune process that involves numerous components depending on the tissue and trigger [198]. The specific role of cytokines in inflammation of the brain is still poorly understood as these ‘danger signals’ [45] are now divided into three different groups: (a) inflammatory cytokines, (b) alarmins, and (c) stressorins, with distinct patterns of secretion and biological properties [199]. Innate immunity of the brain involves primarily microglia [200], which communicate with mast cells [201,202]. Mast cell-derived mediators, such as histamine and tryptase, can activate microglia [203] leading to secretion of pro-inflammatory mediators including interleukins IL-1β, IL-6, and TNF, known to be increased in the brain of children with ASD [204]. Mast cells are found in the brain [205], especially the hypothalamus, thalamus, and third ventricle [206,207]. Mast cells are also found in the pineal, the pituitary, and the thyroid glands [3].
Mast cells can function as the ‘immune gate to the brain’ [208] as they can regulate permeability of the blood–brain barrier (BBB) [209–211] and are involved in neuroinflammation and brain disorders [76,212].
Therefore, a detailed medical history including type, duration and triggers of symptoms, careful and focused physical examination, and specific laboratory tests, is necessary for a proper clinical evaluation. Family history is of great importance because the prevalence of MCD is higher among relatives of MCD patients than would be expected by chance [213].
8. Treatment approaches
8.1. Disclaimer: although treatments are discussed, no endorsement or statement of safety is implied.
Unfortunately, the genetic cause of proliferative mast cell disorders and the effects of the multiple mediators released are reasons for the lack of curative treatments [214–216]. Limiting exposure to triggers, and reduction of symptoms is the main aim of treatment. Inhibiting secretion of mediators, especially cytokines, would be desirable [39],
Common triggers include alcohol, preservatives, spices, heat, cold, drugs [antibiotics, nonsteroidal anti-inflammatory drugs (NSAIDs), opioids, neuromuscular junction blocking agent], radiocontrast media, hymenoptera stings, physical stimuli (pressure, friction), estrogen, and stressful conditions [113]. Reactions to hymenoptera may be the manifestation of nc-MCAD [217], and these patients should carry diphenhydramine (Benadryl, preferably liquid gels) and self-administered intramuscular epinephrine (EpiPen or AnaPen). Patients are usually administrated antihistamines, montelukast, and cromolyn (Table 9).
Table 9.
Treatment Approach for Mast Cell Mediator Disorders
| 1. Antihistamines (cetirizine, diphenhydramine, hydroxyzine) |
| 2. Antihistamines with anti-eosinophilic action (ketotifen, rupatadine) |
| 3. Antihistamine with antiserotonin action (cyproheptadine) |
| 4. Tricyclic antidepressants with combined antihistamine action (doxepin) |
| 5. Flavonoids (luteolin, quercetin of high purity and increased absorption) |
| 6. Antileukotrienes (montelukast) |
| 7. Cromolyn sodium |
| 8. Steroids (methylprednisolone) |
| 9. Epinephrine (EpiPen, AnaPen) |
| 10. Anti-IgE (omalizumab) |
| 11. TK inhibitor (imatinib) |
| 12. Kinase inhibitors for mast cells expressing mutant KIT (mitostaurine, avapritinib) |
8.2. Antihistamines
Prevention of mast cell proliferation and secretion of mast cell mediators is the ultimate goal. Patients are typically treated with histamine (H1,H2)-receptor antagonists (antihistamines) (Table 9). Second generation antihistamines are preferred (loratadine, cetirizine, fexofenadine) because of less sedation and absence of any effects on cardiac excitation [218]. However, before or during invasive procedures (e.g. radiology studies), the first generation diphenhydramine or hydroxyzine would be best. Ketotifen, an H1-receptor antagonist, is also a weak ocular mast cell inhibitor and may be used as a third option [219]. A number of patients who present with eosinophilic esophagitis or gastroenteritis are best treated with rupatadine, which also has anti-platelet activating factor (PAF) and anti-eosinophilic properties actions [220]; rupatadine can also inhibit cultured mast cells [220,221]. The H2-antihistamines (ranitidine, famotidine) are used to block gastric hypersecretion [117]. However, histamine-1 receptor antagonists also have anticholinergic effects [222], and excessive doses may be associated with urinary retention [223] and negatively affect mental status [224,225].
8.3. Mast cell inhibitors
Despite the common use of the term ‘mast cell stabilizers’ [226], there are no such clinically effective drugs. Disodium cromoglycate (cromolyn sodium), known as a ‘mast cell stabilizer,’ reduces mostly GI symptoms, such as abdominal bloating and pain. Cromolyn is administered orally and is divided into doses [117]. Cromolyn causes tachyphylaxis [227], making it necessary to increase the dose from 100 mg tid to 400 mg tid over the course of a year. However, cromolyn is difficult to dissolve, less than 5% is absorbed orally and can cause severe diarrhea in about 15% of patients. Cromolyn can also cause alopecia in about 10% of patients. Recent data have even cast some doubt on the ability of cromolyn to stabilize human mast cells [228], and its beneficial effect on pruritus might be mediated by inhibition of C-fiber peripheral sensory nerves, instead [229].
8.4. Leukotriene receptor antagonists
Increased secretion of leukotriene E4 has been reported in mastocytosis [230]. The cysteinyl-leukotriene receptor antagonist montelukast may be used in patients with refractory symptoms, especially patients with pulmonary involvement (asthma, wheezing).
8.5. Acetylsalicylic acid
Some patients with refractory flushing and elevated urinary 11β-PGF2a have overproduction of prostaglandins [230]. These patients may benefit from acetylsalicylic acid (ASA) for clinical improvement [117,231,232]. However, others may experience hypersensitivity reactions and require desensitization treatment [230,233].
8.6. Omalizumab
Whether patients have elevated serum IgE or not, they may derive significant benefit from frequent use of omalizumab, an anti-IgE humanized murine mAb, approved for allergic asthma, and chronic urticaria. Omalizumab also improves patients in whom IgE levels are not pathologically increased [234]. Omalizumab, reversibly binds to the Fc portion of the free serum IgE molecules, forming circulating immune complexes and reducing IgE binding to FcεRI on mast cells and basophils. As a consequence, FcεRI expression is down-regulated and, surprisingly, reactivity of these cells to other triggers (e.g. pressure) is also reduced [235]. A newer version of omalizumab, mepolizumab, may be equally suitable.
However, omalizumab can also form complement-fixing immune complexes that may lead to serious hypersensitivity reactions including severe ‘serum sickness-like’ reactions and should be administrated in a clinic under supervision. Both the US FDA and the EU EMA have directed inclusion of these potential serious adverse effects in the specialized information accompanying this drug.
Given that omalizumab targets components of the immune system, it may potentially increase the risk of infections or neoplasms.
8.7. Corticosteroids
Corticosteroids could be of great benefit in patients with refractory symptoms especially after resuscitation from an acute episode of anaphylaxis along with the use of subcutaneous epinephrine (EpiPen), and β-agonists [117]. However, there are reported instances of sensitivity to steroids, and their prolonged use is associated with asteoporosis, and depression.
8.8. Venom immunotherapy
Treatment with lifelong venom immunotherapy (VIT) is necessary for patients with MMCAS after hymenoptera stings, especially if they test positive for venom specific IgE [236]. This treatment could have life-threatening side effects and should be undertaken with caution in a clinic under supervision after weighing the risk-benefit for the individual patient.
8.9. Immune Ig
In certain cases, where all else has failed, patients may be treated with immune Ig (ivIg).
8.10. Specific agents
Patients with advanced SM without the D816V KIT mutation may be treated with the tyrosine kinase inhibitor (TKI), imatinib. Another TKI, dasatinib, inhibits D816V KIT expressing mast cells in vitro, but has not been proved successful in clinical trials. A protein kinase C (PKC) inhibitor (previously known as PKC412) approved recently, midostaurin, can inhibit mast cells expressing D816V KIT and was recently approved for advanced forms of clonal proliferative mast cell disorders such as aggressive SM and mast cell leukemia [237]. However, PKC is an important enzyme present in most cells, not only mast cells; hence, the benefit–risk assessment must be carefully weighed. An experimental TKI, avapritinib (formerly known as BLU-285) appears to have even greater ability to reduce mutated KIT mast cells [238].
8.11. Flavonoids
Flavonoids such as quercetin (3′, 4′, 5, 7, 9-pentahydroxyflavone) and luteolin (3′, 4′, 5, 7-tetrahydroxyflavone-NOT lutein, which is a carotenoid) are naturally occurring polyphenolic compounds with potent anti-oxidant, anti-inflammatory, and neuroprotective actions acting by inhibiting multiple targets [239–241]. It was previously shown that flavonols containing more hydroxyl groups have greater anti-oxidant and anti-inflammatory activity [19,209,242], as well as anti-asthmatic effects [218].
8.11.1. Luteolin
Luteolin inhibited mast cells [76,243–245] and decreased histamine secretion in bronchoalveolar lavage fluid from conscious ovalbumin sensitized guinea pigs exposed to aerolized ovalbumin [246]. Luteolin-7-O-glucoside inhibited degranulation and leukotriene production from bone marrow mast cells [247]. Luteolin also inhibited stimulation of human T cells [78], keratinocytes [248], and astrocytes [249]. Luteolin inhibited microglial activation and proliferation [250–254]. A luteolin containing formulation was recently shown to have significant benefit in children with ASD [242,255].
Luteolin also has neuroprotective actions [256–258] and protected against thimerosal-induced inflammatory mediator secretion from human mast cells [244] as well as from methylmercury-induced mitochondrial damage [259]. Luteolin induced synthesis and secretion of neurotrophic factors including nerve growth factor (NGF), Glia-derived growth factor (GDGF), and brain-derived neurotrophic factor (BDNF) [260]. Luteolin is structurally related to 7,8-dihydroflavone, which was shown to have BDNF activity [261], and reduce symptoms in a mouse model of Rett syndrome, most patients with which have symptoms of ASD [262]. Moreover, 3ʹ,4ʹ-dimethoxyflavone was shown to be protective against parthanatos [263].
Both luteolin and tetramethoxyluteolin blocked stimulated intracellular calcium increase [245] and activation of NF-κB in human leukemic HMC-1 mast cells [264]. Moreover, polyphenols have been reported to inhibit mast cell secretion by interfering with the secretory vesicle fusion [265]. The substitution of luteolin’s four hydroxy groups by methoxy groups apparently enhances its anti-inflammatory activity and its benefit in neurodegenerative diseases [266–268].
Flavonoids are difficult to absorb (<10%) after oral administration [269,270] and are extensively metabolized [270–272]. We have been investigating the luteolin structural analog 3ʹ,4ʹ,5,7-tetramethoxyflavone (tetramethoxyluteolin), it is a more potent inhibitor than luteolin on human cultured microglia [251] and mast cells [76]. Methylation of dietary flavones greatly improved metabolic stability and intestinal absorption [273].
Flavonoids are generally considered safe [274–277], and can even protect against chemically induced liver toxicity, a common consequence of many drugs [278]. Moreover, neither luteolin nor tetramethoxyluteolin affects viability of cultured human mast cells [245], T cells [78] or keratinocytes [279]. Luteolin also improved symptoms of ASD [242,280], post-Lyme syndrome [281], and brain fog [139] without any major adverse effects.
However, there should be caution against using flavonoid preparations of unknown source and purity because the cheapest sources are peanut cells and fava beans that could lead to anaphylaxis or hemolytic anemia in G6PD individuals, respectively.
Treatment with luteolin should best be used together with a tetramethoxyluteolin containing skin lotion [282], and requires a few weeks before mast cells are sufficiently inhibited. Luteolin may also be combined with vitamin D3, which has been reported to have anti-allergic actions [283].
9. Summary
A possible treatment approach of patients with mast cell mediator release disorders would be as follows (Table 9): (1) Antihistamines (cetirizine, diphenhydramine, hydroxyzine); (2) antihistamines with anti-eosinophilic action (ketotifen, rupatadine); (3) antihistamine with antiserotonin action (cyproheptadine); (4) tricyclic antidepressants with combined antihistamine action (doxepin); (5) flavonoids (luteolin, quercetin of high purity and increased absorption); (6) antileukotrienes (montelukast); (7) cromolyn sodium; (8) steroids (methylprednisolone); (9) epinephrine (EpiPen, AnaPen); (10) anti-IgE (omalizumab), kinase inhibitors (imatinib, mitostaurine, avapritinib) should be reserved for aggressive SM or mast cell leukemia.
Bone pain is quite common in patients with mast cell disorders and should be addressed [160,216]. Unlike morphine, which is a strong trigger of mast cells, the synthetic opioids tramadol and piritramide (not available in the US) seem to be tolerated. Ketamine and/r propofol, alone or with loaratadine, could be used for anesthesia. Some patients could also tolerate transdermal fentanyl. Low-dose naltrexone, an opioid receptor antagonist, may also be helpful.
Patients with migraine headaches could be helped prophylactically with cyproheptadine or with prochlorperazine, both of which also inhibit human mast cells [284].
10. Expert opinion
Recent reviews stress the role of mast cells in health [285] and disease [215,286], especially their responsiveness to non-allergic triggers [286,287]. There has been considerable progress in defining KIT mutations and developing drugs that block tyrosine kinases (TK) that are involved in mast cell proliferation [288]. However, it has become clear that KIT mutations alone are not able to induce a mast cell proliferative disorder; as a result, the aim of recent research for therapeutic agents is the development of multitargeting drugs. Moreover, our understanding of mast cell stimulation by non-IgE mediated triggers remains poor, and there are still no clinically effective inhibitors of mast cell mediator release. One of the possible reasons for this problem has been the over-reliance on use of rodent mast cells, the results from which have increasingly been shown not to reflect human inflammatory diseases [289].
Moreover, both immortalized and primary normal mast cells cultured from peripheral blood or umbilical cord blood require 8–12 weeks of growth and are ‘immature’ with poorly developed secretory granules [60]. More importantly, mast cells from different tissues are phenotypically very different [290]. For instance, brain mast cells do not express FcɛRI [207], skin mast cells are triggered by morphine, while heart mast cells are not. Moreover, we still do not know differences in cytokine/chemokines release or other mediators. It would be desirable to identify mechanisms and mediators that may contribute to useful physiology as compared to pathological function, and thus stimulate them or inhibit the, respectively. For instance, mast cells have been reported to regulate wound healing in diabetes [291], but contribute to cancer growth and metastasis [292].
Finally, there is the mystery of the presence of mast cells in neuroendocrine organs, such as the pineal, hypothalamus, pituitary, thyroid, and the uterus [3], where the pathophysiologic function of the mast cells remains unknown. These findings may possibly explain our observation that many females with MCMD report increased libido and orgasms [3]. An intriguing relevant observation was the increase of number of mast cells in the habenula of female pigeons during courting [293], and the development of a female ‘phenotype’ in newborn female rats in which mast cells were stimulated [294].
Research efforts should focus on (a) establishing normal mast cell cultures that mimic those in different tissues; (b) investigating the ability of mast cells to respond to different triggers with selective release of mediators; (c) identifying the molecular events involved in mast cell release of inflammatory mediators; (d) studying the role of growth factors other than SCF, such as NGF and platelet-derived growth factor (PDGF); (e) investigating the synergistic actions of triggers, such as SP an IL-33; (f)determining the role of extracellular microvesicles [295]; (g) as well as identifying any innate inhibitors such as the polyamines agmantine (the product of the decarboxylation of arginine) [296] or spermidine and spermine (the products of the decarboxylation of ornithine) [297,298], as well as heparin and chondroitin sulfate [299].
One paper reported on a prototype inhibitory receptor (mast cell functional-associated antigen, MAFA), which selectively regulates FcεRI-stimulated mast cell secretion of IL-1β and CXCL8, but not TNF or MCP-1 [300], but there has not been any apparent further development.
There has been recent interest in the potential mast cell inhibitory ability of the anti-inflammatory cytokine IL-37 [301–303], at least in diseases where mast cells have been implicated [303,304], but direct inhibition of human mast cells has not been reported so far.
Article highlights.
The term MCMD should be used when specific criteria.
Tryptase is specific for mast cells and is used for diagnosis, but is often not secreted and its function is still not known. Measurement of other serum mast cell mediators, especially IL-6 and IL-31, as well as CCL2 and CXCL8, and urine (24 h) histamine metabolites (methyl histamine, MIA), the PGD2 metabolite (11β-PGF2α), and LTE4 should be considered in the absence of any other inflammatory conditions.
Screening tissue sections for mast cells should combine immunohistochemistry for CD117 (identifies all mast cells) and tryptase (identifies degranulated mast cells).
Single cell laser microcapture and qRT-PCR should be used to identify mast cell phenotype variability and mediator synthesis/release in situ.
Special attention should be placed on the timing of measurement of mast cell mediators and patient symptoms as mast cells have been shown to be regulated by a circadian clock.
There is no curative treatment for MCMDs at the present time. Prevention of stimulation of mast cells and reduction of the effects of their mediators are necessary. Step-wise increase in treatment is recommended as indicated and tolerated.
Development of effective mast cell inhibitors should be a top priority. Recent promising results from the use of tetramethoxyluteolin and IL-37 should be expanded in appropriate formulations.
Funding
Aspects of the work discussed here were supported in part by US National Institutes of Health (NIH) grants NS055681; DK062861; AR47652; NS66205; NS71361, from Pfizer and the National Psoriasis Foundation and from Theta Biomedical Consulting and Development Co., Inc. (Brookline, MA).
Footnotes
Declaration of interest
TC Theoharides is on the Scientific Advisory Board of The Mastocytosis Society (TMS, www.tmsforacure.org) TC Theoharides is the Scientific Director of Algonot, LLC, which has developed flavonoid containing dietary supplements. TC Theoharides is the inventor of US patent nos. 6,635,625; 6,641,806; 6,645,482; 6,689,748; 6,984,667; 7,906,153; 8,268,365; 9,050,275; and 9,76,146, as well as EPO 1365777 covering the use of proteoglycans and flavonoids for the treatment of mast cell-related and inflammatory diseases. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed. No writing assistance was utilized in the production of this manuscript.
Reviewer disclosures
Peer reviewers on this manuscript have no relevant financial or other relationships to disclose.
References
Papers of special note have been highlighted as either of interest (•) or of considerable interest (••) to readers.
- 1.Chen CC, Grimbaldeston MA, Tsai M, et al. Identification of mast cell progenitors in adult mice. Proc Natl Acad Sci U S A. 2005;102:11408–11413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Theoharides TC, Valent P, Akin C. Mast cells, mastocytosis, and related disorders. N Engl J Med. 2015;373:163–172. [DOI] [PubMed] [Google Scholar]
- 3.•.Theoharides TC. Neuroendocrinology of mast cells: challenges and controversies. Exp Dermatol. 2017;26:751–759. This paper reviews evidence of the responsiveness of mast cells to neuroendocrine triggers and their potential role in related diseases in the absence of allergies or MCMD. [DOI] [PubMed] [Google Scholar]
- 4.••.Cheng LE, Hartmann K, Roers A, et al. Perivascular mast cells dynamically probe cutaneous blood vessels to capture immunoglobulin E. Immunity. 2013;38(1):166–175. This is the first report of demonstrating mast cells sensing the blood vessels and capturing circulating IgE. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Galli SJ, Tsai M, Piliponsky AM. The development of allergic inflammation. Nature. 2008;454:445–454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Toniato E, Frydas I, Robuffo I, et al. Activation and inhibition of adaptive immune response mediated by mast cells. J Biol Regul Homeost Agents. 2017;31:543–548. [PubMed] [Google Scholar]
- 7.Avila M, Gonzalez-Espinosa C. Signaling through Toll-like receptor 4 and mast cell-dependent innate immunity responses. IUBMB Life. 2011;63:873–880. [DOI] [PubMed] [Google Scholar]
- 8.Cardamone C, Parente R, Feo GD, et al. Mast cells as effector cells of innate immunity and regulators of adaptive immunity. Immunol Lett. 2016;178:10–14. [DOI] [PubMed] [Google Scholar]
- 9.Forsythe P Microbes taming mast cells: implications for allergic inflammation and beyond. Eur J Pharmacol. 2016;778:169–175. [DOI] [PubMed] [Google Scholar]
- 10.Gong J, Yang NS, Croft M, et al. The antigen presentation function of bone marrow-derived mast cells is spatiotemporally restricted to a subset expressing high levels of cell surface FcepsilonRI and MHC II. BMC Immunol. 2010;11:34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.•.Carroll-Portillo A, Cannon JL, Te RJ, et al. Mast cells and dendritic cells form synapses that facilitate antigen transfer for T cell activation. J Cell Biol. 2015;210:851–864. Important finding of direct mast cell-dendritic contact facilitating antigen transfer for T cell activation. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bulfone-Paus S, Bahri R. Mast cells as regulators of T cell responses. Front Immunol. 2015;6:394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ishii T, Wang J, Zhang W, et al. Pivotal role of mast cells in pruritogenesis in patients with myeloproliferative disorders. Blood. 2009;113:5942–5950. [DOI] [PubMed] [Google Scholar]
- 14.Mekori YA, Hershko AY, Frossi B, et al. Integrating innate and adaptive immune cells: mast cells as crossroads between regulatory and effector B and T cells. Eur J Pharmacol. 2015. [DOI] [PubMed] [Google Scholar]
- 15.Rodriguez Cetina BH, Heinbokel T, Uehara H, et al. Mast cells regulate CD4(+) T-cell differentiation in the absence of antigen presentation. J Allergy Clin Immunol. 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Christy AL, Brown MA. The multitasking mast cell: positive and negative roles in the progression of autoimmunity. J Immunol. 2007;179:2673–2679. [DOI] [PubMed] [Google Scholar]
- 17.Theoharides TC, Alysandratos KD, Angelidou A, et al. Mast cells and inflammation. Biochim Biophys Acta. 2012;1822:21–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sismanopoulos N, Delivanis DA, Mavrommati D, et al. Do mast cells link obesity and asthma? Allergy. 2013;68:8–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mukai K, Tsai M, Saito H, et al. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol Rev. 2018;282:121–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Galli SJ, Grimbaldeston M, Tsai M. Immunomodulatory mast cells: negative, as well as positive, regulators of immunity. Nat Rev Immunol. 2008;8:478–486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cao J, Papadopoulou N, Kempuraj D, et al. Human mast cells express corticotropin-releasing hormone (CRH) receptors and CRH leads to selective secretion of vascular endothelial growth factor. J Immunol. 2005;174:7665–7675. [DOI] [PubMed] [Google Scholar]
- 22.Lazarus LH, Perrin MH, Brown MR, et al. Verification of both the sequence and confromational specificity of neurotensin in binding to mast cells. Biochem Biophys Res Commun. 1977;76:1079–1085. [DOI] [PubMed] [Google Scholar]
- 23.Donelan J, Boucher W, Papadopoulou N, et al. Corticotropin-releasing hormone induces skin vascular permeability through a neurotensin-dependent process. Proc Natl Acad Sci USA. 2006;103:7759–7764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mustain WC, Rychahou PG, Evers BM. The role of neurotensin in physiologic and pathologic processes. Curr Opin Endocrinol Diabetes Obes. 2011;18:75–82. [DOI] [PubMed] [Google Scholar]
- 25.Caceda R, Kinkead B, Nemeroff CB. Neurotensin: role in psychiatric and neurological diseases. Peptides. 2006;27:2385–2404. [DOI] [PubMed] [Google Scholar]
- 26.Mashaghi A, Marmalidou A, Tehrani M, et al. Neuropeptide substance P and the immune response. Cell Mol Life Sci. 2016;73:4249–4264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.O’Connor TM, O’Connell J, O’Brien DI, et al. The role of substance P in inflammatory disease. J Cell Physiol. 2004;201:167–180. [DOI] [PubMed] [Google Scholar]
- 28.Hokfelt T, Pernow B, Wahren J. Substance P: a pioneer amongst neuropeptides. J Intern Med. 2001;249:27–40. [DOI] [PubMed] [Google Scholar]
- 29.Douglas SD, Leeman SE. Neurokinin-1 receptor: functional significance in the immune system in reference to selected infections and inflammation. Ann N Y Acad Sci. 2011;1217:83–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.•.McNeil BD, Pundir P, Meeker S, et al. Identification of a mast-cell-specific receptor crucial for pseudo-allergic drug reactions. Nature. 2015;519:237–241. Identification of a low affinity receptor that appears to mediate stimulation of mast cells by cationic molecules. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Theoharides TC, Kempuraj D, Tagen M, et al. Differential release of mast cell mediators and the pathogenesis of inflammation. Immunol Rev. 2007;217:65–78. [DOI] [PubMed] [Google Scholar]
- 32.Zhang B, Weng Z, Sismanopoulos N, et al. Mitochondria distinguish granule-stored from de novo synthesized tumor necrosis factor secretion in human mast cells. Int Arch Allergy Immunol. 2012;159:23–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Askenase PW. Mast cells and the mediation of T-cell recruitment in arthritis. N Engl J Med. 2005;349:1294. [DOI] [PubMed] [Google Scholar]
- 34.Zhang B, Alysandratos KD, Angelidou A, et al. Human mast cell degranulation and preformed TNF secretion require mitochondrial translocation to exocytosis sites: relevance to atopic dermatitis. J Allergy Clin Immunol. 2011;127:1522–1531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gordon JR, Galli SJ. Mast cells as a source of both preformed and immunologically inducible TNF-α/cachectin. Nature. 1990;346:274–276. [DOI] [PubMed] [Google Scholar]
- 36.Picard M, Giavina-Bianchi P, Mezzano V, et al. Expanding spectrum of mast cell activation disorders: monoclonal and idiopathic mast cell activation syndromes. Clin Ther. 2013;35:548–562. [DOI] [PubMed] [Google Scholar]
- 37.••.Theoharides TC, Bondy PK, Tsakalos ND, et al. Differential release of serotonin and histamine from mast cells. Nature. 1982;297:229–231. First report of the ability of mast cells to release mediators without degranulation. [DOI] [PubMed] [Google Scholar]
- 38.Theoharides TC, Cochrane DE. Critical role of mast cells in inflammatory diseases and the effect of acute stress. J Neuroimmunol. 2004;146:1–12. [DOI] [PubMed] [Google Scholar]
- 39.Caraffa A, Conti C, Ovidio D, et al. New concepts in neuroinflammation: mast cells pro-inflammatory and anti-inflammatory cytokine mediators. J Biol Regul Homeost Agents. 2018;32:449–454. [PubMed] [Google Scholar]
- 40.Conti P, Caraffa A, Kritas SK, et al. Mast cell, pro-inflammatory and anti-inflammatory: jekyll and Hyde, the story continues. J Biol Regul Homeost Agents. 2017;31:263–267. [PubMed] [Google Scholar]
- 41.Salamon P, Shoham NG, Gavrieli R, et al. Human mast cells release interleukin-8 and induce neutrophil chemotaxis on contact with activated T cells. Allergy. 2005;60:1316–1319. [DOI] [PubMed] [Google Scholar]
- 42.•.Kempuraj D, Papadopoulou NG, Lytinas M, et al. Corticotropin-releasing hormone and its structurally related urocortin are synthesized and secreted by human mast cells. Endocrinology. 2004;145:43–48. First report that human mast cells can synthesize and release the stress-related peptide, corticotropin-releasing hormone (CRH, or factor, CRF). [DOI] [PubMed] [Google Scholar]
- 43.Theoharides TC, Donelan JM, Papadopoulou N, et al. Mast cells as targets of corticotropin-releasing factor and related peptides. Trends Pharmacol Sci. 2004;25:563–568. [DOI] [PubMed] [Google Scholar]
- 44.Petra AI, Tsilioni I, Taracanova A, et al. Interleukin 33 and interleukin 4 regulate interleukin 31 gene expression and secretion from human laboratory of allergic diseases 2 mast cells stimulated by substance P and/or immunoglobulin E. Allergy Asthma Proc. 2018;39(2):153–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Theoharides TC. Danger signals and inflammation. Clin Ther. 2016;38:996–999. [DOI] [PubMed] [Google Scholar]
- 46.Zhang B, Asadi S, Weng Z, et al. Stimulated human mast cells secrete mitochondrial components that have autocrine and paracrine inflammatory actions. PloS One. 2012;7:e49767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Collins LV, Hajizadeh S, Holme E, et al. Endogenously oxidized mitochondrial DNA induces in vivo and in vitro inflammatory responses. J Leukoc Biol. 2004;75:995–1000. [DOI] [PubMed] [Google Scholar]
- 48.Marques PE, Amaral SS, Pires DA, et al. Chemokines and mitochondrial products activate neutrophils to amplify organ injury during mouse acute liver failure. Hepatology. 2012;56:1971–1982. [DOI] [PubMed] [Google Scholar]
- 49.Sun S, Sursal T, Adibnia Y, et al. Mitochondrial DAMPs increase endothelial permeability through neutrophil dependent and independent pathways. PloS One. 2013;8:e59989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Theoharides TC, Asadi S, Panagiotidou S, et al. The “missing link” in autoimmunity and autism: extracellular mitochondrial components secreted from activated live mast cells. Autoimmun Rev. 2013;12:1136–1142. [DOI] [PubMed] [Google Scholar]
- 51.Asadi S, Theoharides TC. Corticotropin-releasing hormone and extracellular mitochondria augment IgE-stimulated human mast-cell vascular endothelial growth factor release, which is inhibited by luteolin. J Neuroinflam. 2012;9:85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lauritzen KH, Moldestad O, Eide L, et al. Mitochondrial DNA toxicity in forebrain neurons causes apoptosis, neurodegeneration, and impaired behavior. Mol Cell Biol. 2010;30:1357–1367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zhang B, Angelidou A, Alysandratos KD, et al. Mitochondrial DNA and anti-mitochondrial antibodies in serum of autistic children. J Neuroinflammation. 2010;7:80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Moulin D, Donze O, Talabot-Ayer D, et al. Interleukin (IL)-33 induces the release of pro-inflammatory mediators by mast cells. Cytokine. 2007;40:216–225. [DOI] [PubMed] [Google Scholar]
- 55.Saluja R, Ketelaar ME, Hawro T, et al. The role of the IL-33/IL-1RL1 axis in mast cell and basophil activation in allergic disorders. Mol Immunol. 2015;63:80–85. [DOI] [PubMed] [Google Scholar]
- 56.Theoharides TC, Petra AI, Taracanova A, et al. Targeting IL-33 in autoimmunity and inflammation. J Pharmacol Exp Ther. 2015;354:24–31. [DOI] [PubMed] [Google Scholar]
- 57.Silver MR, Margulis A, Wood N, et al. IL-33 synergizes with IgE-dependent and IgE-independent agents to promote mast cell and basophil activation. Inflamm Res. 2010;59:207–218. [DOI] [PubMed] [Google Scholar]
- 58.Kumar S, Khodoun M, Kettleson EM, et al. Glyphosate-rich air samples induce IL-33, TSLP and generate IL-13 dependent airway inflammation. Toxicology. 2014;325:42–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Theoharides TC, Zhang B, Kempuraj D, et al. IL-33 augments substance P-induced VEGF secretion from human mast cells and is increased in psoriatic skin. Proc Natl Acad Sci USA. 2010;107:4448–4453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.••.Taracanova A, Alevizos M, Karagkouni A, et al. SP and IL-33 together markedly enhance TNF synthesis and secretion from human mast cells mediated by the interaction of their receptors. Proc Natl Acad Sci U S A. 2017;114:E4002–E4009. First report that stimulation of human mast cells by the neuropeptide substance P and the cytokine IL-33 leads to strong synergistic expression of TNF, which is inhibited by the natural flavonoid methoxyluteolin. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.••.Taracanova A, Tsilioni I, Conti P, et al. Substance P and IL-33 administered together stimulate a marked secretion of IL-1beta from human mast cells, inhibited by methoxyluteolin. Proc Natl Acad Sci U S A. 2018. First report that stimulation of human mast cells by the neuropeptide substance P and the cytokine IL-33 leads to strong synergistic expression of IL-1β, which is inhibited by the natural flavonoid methoxyluteolin. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hsu CL, Neilsen CV, Bryce PJ. IL-33 is produced by mast cells and regulates IgE-dependent inflammation. PLoS ONE. 2010;5:e11944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Tung HY, Plunkett B, Huang SK, et al. Murine mast cells secrete and respond to interleukin-33. J Interferon Cytokine Res. 2014;34:141–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sumpter TL, Ho CH, Pleet AR, et al. Autocrine hemokinin-1 functions as an endogenous adjuvant for IgE-mediated mast cell inflammatory responses. J Allergy Clin Immunol. 2015;135:1019–1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Morita H, Nakae S, Saito H, et al. IL-33 in clinical practice: size matters? J Allergy Clin Immunol. 2017;140:381–383. [DOI] [PubMed] [Google Scholar]
- 66.•.Kandere-Grzybowska K, Letourneau R, Kempuraj D, et al. IL-1 induces vesicular secretion of IL-6 without degranulation from human mast cells. J Immunol. 2003;171:4830–4836. First report that human mast cells can release cytokines via vesicular release unrelated to secretory granules. [DOI] [PubMed] [Google Scholar]
- 67.Noordenbos T, Blijdorp I, Chen S, et al. Human mast cells capture, store, and release bioactive, exogenous IL-17A. J Leukoc Biol. 2016;100:453–462. [DOI] [PubMed] [Google Scholar]
- 68.Theoharides TC, Kalogeromitros D. The critical role of mast cells in allergy and inflammation. Ann N Y Acad Sci. 2006;1088:78–99. [DOI] [PubMed] [Google Scholar]
- 69.Wernersson S, Pejler G. Mast cell secretory granules: armed for battle. Nat Rev Immunol. 2014;14:478–494. [DOI] [PubMed] [Google Scholar]
- 70.Xu H, Bin NR, Sugita S. Diverse exocytic pathways for mast cell mediators. Biochem Soc Trans. 2018;46:235–247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Gilfillan AM, Tkaczyk C. Integrated signalling pathways for mast-cell activation. Nat Rev Immunol. 2006;6:218–230. [DOI] [PubMed] [Google Scholar]
- 72.Gaudenzio N, Sibilano R, Marichal T, et al. Different activation signals induce distinct mast cell degranulation strategies. J Clin Invest. 2016;126:3981–3998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Sibilano R, Frossi B, Pucillo CE. Mast cell activation: a complex interplay of positive and negative signaling pathways. Eur J Immunol. 2014;44:2558–2566. [DOI] [PubMed] [Google Scholar]
- 74.Crivellato E, Nico B, Gallo VP, et al. Cell secretion mediated by granule-associated vesicle transport: a glimpse at evolution. Anat Rec (Hoboken) 2010;293:1115–1124. [DOI] [PubMed] [Google Scholar]
- 75.Smrz D, Kim MS, Zhang S, et al. mTORC1 and mTORC2 differentially regulate homeostasis of neoplastic and non-neoplastic human mast cells. Blood. 2011;118:6803–6813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.•.Patel AB, Theoharides TC. Methoxyluteolin inhibits neuropeptide-stimulated proinflammatory mediator release via mTOR activation from human mast cells. J Pharmacol Exp Ther. 2017;361:462–471. First report that stimulation of human mast cells by neuropeptides is mediated by mTOR activation that is inhibited by the natural flavonoid methoxyluteolin. [DOI] [PubMed] [Google Scholar]
- 77.Nakae S, Suto H, Kakurai M, et al. Mast cells enhance T cell activation: importance of mast cell-derived TNF. Proc Natl Acad Sci U S A. 2005;102:6467–6472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kempuraj D, Tagen M, Iliopoulou BP, et al. Luteolin inhibits myelin basic protein-induced human mast cell activation and mast cell dependent stimulation of Jurkat T cells. Br J Pharmacol. 2008;155:1076–1084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Nakamura Y, Nakano N, Ishimaru K, et al. Inhibition of IgE-mediated allergic reactions by pharmacologically targeting the circadian clock. J Allergy Clin Immunol. 2016;137:1226–1235. [DOI] [PubMed] [Google Scholar]
- 80.••.Nakao A, Nakamura Y, Shibata S. The circadian clock functions as a potent regulator of allergic reaction. Allergy. 2015;70:467–473. Important finding that stimulation of mast cells and release of mediators may be regulated by circadian rhythms that may help with dignosis and pathogenesis of allergic diseases. [DOI] [PubMed] [Google Scholar]
- 81.Hatziagelaki E, Adamaki M, Dimitriadis G, et al. Myalgic encephalomyelitis/chronic fatigue syndrome-metabolic diseases or disturbed homeostasis? Jpet 2018;In press. [DOI] [PubMed] [Google Scholar]
- 82.Theoharides TC, Tsilioni I, Arbetman L, et al. Fibromyalgia, a syndrome in search of pathogenesis and therapy. J Pharmacol Exp Ther. 2015;355:255–263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Theoharides TC, Sismanopoulos N, Delivanis DA, et al. Mast cells squeeze the heart and stretch the gird: their role in atherosclerosis and obesity. Trends Pharmacol Sci. 2011;32:534–542. [DOI] [PubMed] [Google Scholar]
- 84.Alevizos M, Karagkouni A, Panagiotidou S, et al. Stress triggers coronary mast cells leading to cardiac events. Ann Allergy Asthma Immunol. 2013;112:309–316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Metcalfe DD, Schwartz LB. Assessing anaphylactic risk? Consider mast cell clonality. J Allergy Clin Immunol. 2009;123:687–688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Bonadonna P, Perbellini O, Passalacqua G, et al. Clonal mast cell disorders in patients with systemic reactions to Hymenoptera stings and increased serum tryptase levels. J Allergy Clin Immunol. 2009;123:680–686. [DOI] [PubMed] [Google Scholar]
- 87.Niedoszytko M, de MJ, van Doormaal JJ, et al. Mastocytosis and insect venom allergy: diagnosis, safety and efficacy of venom immunotherapy. Allergy. 2009;64:1237–1245. [DOI] [PubMed] [Google Scholar]
- 88.Bonadonna P, Zanotti R, Muller U. Mastocytosis and insect venom allergy. Curr Opin Allergy Clin Immunol. 2010;10:347–353. [DOI] [PubMed] [Google Scholar]
- 89.Wimazal FGPSPSWVP. Severe life-threatening or disabling anaphylaxis in patients with systemic mastocytosis: a single-center experience. Int Arch Allergy Immunol. 2012;417–424. [DOI] [PubMed] [Google Scholar]
- 90.Lieberman P, Nicklas RA, Oppenheimer J, et al. The diagnosis and management of anaphylaxis practice parameter: 2010 update. J Allergy Clin Immunol. 2010;126:477–480. [DOI] [PubMed] [Google Scholar]
- 91.Kritas SK, Saggini A, Varvara G, et al. Mast cell involvement in rheumatoid arthritis. J Biol Regul Homeost Agents. 2013;27:655–660. [PubMed] [Google Scholar]
- 92.Schwartz LB. Mast cells and basophils. Clin Allergy Immunol. 2002;16:3–42. [PubMed] [Google Scholar]
- 93.Hauswirth AW, Natter S, Ghannadan M, et al. Recombinant allergens promote expression of CD203c on basophils in sensitized individuals. J Allergy Clin Immunol. 2002;110:102–109. [DOI] [PubMed] [Google Scholar]
- 94.Valent P, Hauswirth AW, Natter S, et al. Assays for measuring in vitro basophil activation induced by recombinant allergens. Methods. 2004;32:265–270. [DOI] [PubMed] [Google Scholar]
- 95.Hauswirth AW, Escribano L, Prados A, et al. CD203c is overexpressed on neoplastic mast cells in systemic mastocytosis and is upregulated upon IgE receptor cross-linking. Int J Immunopathol Pharmacol. 2008;21:797–806. [DOI] [PubMed] [Google Scholar]
- 96.Wilcock A, Bahri R, Bulfone-Paus S, et al. Mast cell disorders: from infancy to maturity. Allergy. 2018. [DOI] [PubMed] [Google Scholar]
- 97.Metcalfe DD, Mekori YA. Pathogenesis and pathology of mastocytosis. Annu Rev Pathol. 2017;12:487–514. [DOI] [PubMed] [Google Scholar]
- 98.Valent P, Akin C, Arock M, et al. Definitions, criteria and global classification of mast cell disorders with special reference to mast cell activation syndromes: a consensus proposal. Int Arch Allergy Immunol. 2012;157:215–225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Akin C, Valent P, Metcalfe DD. Mast cell activation syndrome: proposed diagnostic criteria. J Allergy Clin Immunol. 2010;126:1099–1104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Molderings GJ. The genetic basis of mast cell activation disease - looking through a glass darkly. Crit Rev Oncol Hematol. 2015;93:75–89. [DOI] [PubMed] [Google Scholar]
- 101.Pardanani A Systemic mastocytosis in adults: 2019 update on diagnosis, risk stratification and management. Am J Hematol. 2019;94:363–377. [DOI] [PubMed] [Google Scholar]
- 102.Horny HP, Sotlar K, Valent P. Evaluation of mast cell activation syndromes: impact of pathology and immunohistology. Int Arch Allergy Immunol. 2012;159:1–5. [DOI] [PubMed] [Google Scholar]
- 103.Butterfield JH, Li CY. Bone marrow biopsies for the diagnosis of systemic mastocytosis: is one biopsy sufficient?. Am J Clin Pathol. 2004;121:264–267. [DOI] [PubMed] [Google Scholar]
- 104.Valent P, Akin C, Escribano L, et al. Standards and standardization in mastocytosis: consensus statements on diagnostics, treatment recommendations and response criteria. Eur J Clin Invest. 2007;37:435–453. [DOI] [PubMed] [Google Scholar]
- 105.Lindner PS, Pardanani B, Angadi C, et al. Acute nonlymphocytic leukemia in systemic mastocytosis with biclonal gammopathy. J Allergy Clin Immunol. 1992;90:410–412. [DOI] [PubMed] [Google Scholar]
- 106.Sonneck K, Florian S, Mullauer L, et al. Diagnostic and subdiagnostic accumulation of mast cells in the bone marrow of patients with anaphylaxis: monoclonal mast cell activation syndrome. Int Arch Allergy Immunol. 2006;142:158–164. [DOI] [PubMed] [Google Scholar]
- 107.Valent P, Horny HP, Escribano L, et al. Diagnostic criteria and classification of mastocytosis: a consensus proposal. Leuk Res. 2001;25:603–625. [DOI] [PubMed] [Google Scholar]
- 108.Alvarez-Twose I, Zanotti R, Gonzalez-de-Olano D, et al. Nonaggressive systemic mastocytosis (SM) without skin lesions associated with insect-induced anaphylaxis shows unique features versus other indolent SM. J Allergy Clin Immunol. 2013. [DOI] [PubMed] [Google Scholar]
- 109.Wang SA, Hutchinson L, Tang G, et al. Systemic mastocytosis with associated clonal hematological non-mast cell lineage disease: clinical significance and comparison of chomosomal abnormalities in SM and AHNMD components. Am J Hematol. 2013;88:219–224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Furitsu T, Tsujimura T, Tono T, et al. Identification of mutations in the coding sequence of the proto-oncogene c-kit in a human mast cell leukemia cell line causing ligand-independent activation of c-kit product. J Clin Invest. 1993;92:1736–1744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Gagari E, Tsai M, Lantz CS, et al. Differential release of mast cell interleukin-6 via c-kit. Blood. 1997;89:2654–2663. [PubMed] [Google Scholar]
- 112.Akin C, Scott LM, Kocabas CN, et al. Demonstration of an aberrant mast-cell population with clonal markers in a subset of patients with “idiopathic” anaphylaxis. Blood. 2007;110:2331–2333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Hamilton MJ, Hornick JL, Akin C, et al. Mast cell activation syndrome: a newly recognized disorder with systemic clinical manifestations. J Allergy Clin Immunol. 2011;128:147–152. [DOI] [PubMed] [Google Scholar]
- 114.Valent P, Horny HP, Triggiani M, et al. Clinical and laboratory parameters of mast cell activation as basis for the formulation of diagnostic criteria. Int Arch Allergy Immunol. 2011;156:119–127. [DOI] [PubMed] [Google Scholar]
- 115.Pardanani A, Brockman SR, Paternoster SF, et al. FIP1L1-PDGFRA fusion: prevalence and clinicopathologic correlates in 89 consecutive patients with moderate to severe eosinophilia. Blood. 2004;104:3038–3045. [DOI] [PubMed] [Google Scholar]
- 116.Matito A, Azana JM, Torrelo A, et al. Cutaneous mastocytosis in adults and children: new classification and prognostic factors. Immunol Allergy Clin North Am. 2018;38:351–363. [DOI] [PubMed] [Google Scholar]
- 117.Castells M, Metcalfe DD, Escribano L. Diagnosis and treatment of cutaneous mastocytosis in children: practical recommendations. Am J Clin Dermatol. 2011;12:259–270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Cardet JC, Castells MC, Hamilton MJ. Immunology and clinical manifestations of non-clonal mast cell activation syndrome. Curr Allergy Asthma Rep. 2013;13:10–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Alvarez-Twose I, Vano-Galvan S, Sanchez-Munoz L, et al. Increased serum baseline tryptase levels and extensive skin involvement are predictors for the severity of mast cell activation episodes in children with mastocytosis. Allergy. 2012;67:813–821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Barnes M, Van L, DeLong L, et al. Severity of cutaneous findings predict the presence of systemic symptoms in pediatric maculopapular cutaneous mastocytosis. Pediatr Dermatol. 2014;31:271–275. [DOI] [PubMed] [Google Scholar]
- 121.Hosking AM, Makdisi J, Ortenzio F, et al. Diffuse cutaneous mastocytosis: case report and literature review. Pediatr Dermatol. 2018;35:e348–e352. [DOI] [PubMed] [Google Scholar]
- 122.Carter MC, Bai Y, Ruiz-Esteves KN, et al. Detection of KIT D816V in peripheral blood of children with manifestations of cutaneous mastocytosis suggests systemic disease. Br J Haematol. 2018;183:775–782. [DOI] [PubMed] [Google Scholar]
- 123.Horigome K, Bullock ED, Johnson EM Jr. Effects of nerve growth factor on rat peritoneal mast cells. J Biol Chem. 1994;169:2695–2702. [PubMed] [Google Scholar]
- 124.Conti P, Pang X, Boucher W, et al. Impact of Rantes and MCP-1 chemokines on in vivo basophilic mast cell recruitment in rat skin injection model and their role in modifying the protein and mRNA levels for histidine decarboxylase. Blood. 1997;89:4120–4127. [PubMed] [Google Scholar]
- 125.Ito N, Sugawara K, Bodo E, et al. Corticotropin-releasing hormone stimulates the in situ generation of mast cells from precursors in the human hair follicle mesenchyme. J Invest Dermatol. 2010;130:995–1004. [DOI] [PubMed] [Google Scholar]
- 126.Kalogeromitros D, Gregoriou S, Makris M, et al. Secondary anetoderma associated with mastocytosis. Int Arch Allergy Immunol. 2007;142:86–88. [DOI] [PubMed] [Google Scholar]
- 127.Bonadonna P, Scaffidi L. Hymenoptera anaphylaxis as a clonal mast cell disorder. Immunol Allergy Clin North Am. 2018;38:455–468. [DOI] [PubMed] [Google Scholar]
- 128.Worobec AS, Semere T, Nagata H, et al. Clinical correlates of the presence of the Asp816Val c-kit mutation in the peripheral blood mononuclear cells of patients with mastocytosis. Cancer. 1998;83:2120–2129. [DOI] [PubMed] [Google Scholar]
- 129.Mousli M, Bueb J-L, Bronner C, et al. G protein activation: a receptor-independent mode of action for cationic amphiphilic neuropeptides and venom peptides. T I P S. 1990;11:358–362. [DOI] [PubMed] [Google Scholar]
- 130.Woolhiser MR, Brockow K, Metcalfe DD. Activation of human mast cells by aggregated IgG through FcγRI: additive effects of C3a. Clin Immunol. 2004;110:172–180. [DOI] [PubMed] [Google Scholar]
- 131.Abraham SN, St John AL. Mast cell-orchestrated immunity to pathogens. Nat Rev Immunol. 2010;10:440–452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Shih AR, Deshpande V, Ferry JA, et al. Clinicopathological characteristics of systemic mastocytosis in the intestine. Histopathology. 2016;69:1021–1027. [DOI] [PubMed] [Google Scholar]
- 133.Doyle LA, Sepehr GJ, Hamilton MJ, et al. A clinicopathologic study of 24 cases of systemic mastocytosis involving the gastrointestinal tract and assessment of mucosal mast cell density in irritable bowel syndrome and asymptomatic patients. Am J Surg Pathol. 2014;38:832–843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Akin C Mast Cell Activation Disorders. J Allergy Clin Immunol Pract. 2014;2:252–257. [DOI] [PubMed] [Google Scholar]
- 135.Petra AI, Panagiotidou S, Stewart JM, et al. Spectrum of mast cell activation disorders. Expert Rev Clin Immunol. 2014;10:729–739. [DOI] [PubMed] [Google Scholar]
- 136.Afrin LB, Self S, Menk J, et al. Characterization of Mast Cell Activation Syndrome. Am J Med Sci. 2017;353:207–215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Valent P, Akin C, Bonadonna P, et al. Mast cell activation syndrome: importance of consensus criteria and call for research. J Allergy Clin Immunol. 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Valent P, Akin C, Bonadonna P, et al. Proposed diagnostic algorithm for patients with suspected mast cell activation syndrome. J Allergy Clin Immunol Pract. 2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Theoharides TC, Stewart JM, Hatziagelaki E, et al. Brain “fog,” inflammation and obesity: key aspects of 2 neuropsychiatric disorders improved by luteolin. Front Neurosci. 2015;9:225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Ghosh D, Bernstein JA. Systemic and localized seminal plasma hypersensitivity patients exhibit divergent immunologic characteristics. J Allergy Clin Immunol. 2014;134: 969–972. [DOI] [PubMed] [Google Scholar]
- 141.Russell N, Jennings S, Jennings B, et al. The mastocytosis society survey on mast cell disorders: part 2-patient clinical experiences and beyond. J Allergy Clin Immunol Pract. 2018. [DOI] [PubMed] [Google Scholar]
- 142.Jennings S, Russell N, Jennings B, et al. The mastocytosis society survey on mast cell disorders: patient experiences and perceptions. J Allergy Clin Immunol Pract. 2014;2:70–76. [DOI] [PubMed] [Google Scholar]
- 143.Moura DS, Georgin-Lavialle S, Gaillard R, et al. Neuropsychological features of adult mastocytosis. Immunol Allergy Clin North Am. 2014;34:407–422. [DOI] [PubMed] [Google Scholar]
- 144.Moura DS, Sultan S, Georgin-Lavialle S, et al. Evidence for cognitive impairment in mastocytosis: prevalence, features and correlations to depression. PLoS ONE. 2012;7:e39468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Afrin LB, Pohlau D, Raithel M, et al. Mast cell activation disease: an underappreciated cause of neurologic and psychiatric symptoms and diseases. Brain Behav Immun. 2015;50:314–321. [DOI] [PubMed] [Google Scholar]
- 146.Theoharides TC, Kempuraj D, Marchand J, et al. Urticaria pigmentosa associated with acute stress and lesional skin mast cell expression of CRF-R1. Clin Exp Dermatol. 2008;34:e163–e166. [DOI] [PubMed] [Google Scholar]
- 147.Theoharides TC, Petra AI, Stewart JM, et al. High serum corticotropin-releasing hormone (CRH) and bone marrow mast cell CRH receptor expression in a mastocytosis patient. J Allergy Clin Immunol. 2014;134:1197–1199. [DOI] [PubMed] [Google Scholar]
- 148.Theoharides TC. Mold and Immunity. Clin Ther. 2018;40:882–884. [DOI] [PubMed] [Google Scholar]
- 149.Ratnaseelan AM, Tsilioni I, Theoharides TC. Effects of mycotoxins on neuropsychiatric symptoms and immune processes. Clin Ther. 2018;40:903–917. [DOI] [PubMed] [Google Scholar]
- 150.Romo-Lozano Y, Hernandez-Hernandez F, Salinas E. Sporothrix schenckii yeasts induce ERK pathway activation and secretion of IL-6 and TNF-alpha in rat mast cells, but no degranulation. Med Mycol. 2014;52:862–868. [DOI] [PubMed] [Google Scholar]
- 151.Talkington J, Nickell SP. Borrelia burgdorferi spirochetes induce mast cell activation and cytokine release. Infect Immun. 1999;67:1107–1115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Maintz L, Novak N. Histamine and histamine intolerance. Am J Clin Nutr. 2007;85:1185–1196. [DOI] [PubMed] [Google Scholar]
- 153.Bulfone-Paus S, Nilsson G, Draber P, et al. Negative signals in mast cell activation. Trends Immunol. 2017;38:657–667. [DOI] [PubMed] [Google Scholar]
- 154.Hanjra P, Lee CR, Maric I, et al. Chromogranin A is not a biomarker of mastocytosis. J Allergy Clin Immunol Pract. 2018;6:687–689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Theoharides TC. Atopic conditions in search of pathogenesis and therapy. Clin Ther. 2013;35:544–547. [DOI] [PubMed] [Google Scholar]
- 156.Theoharidess TC. Irritable bowel syndrome and ulcerative colitis: mast cell numbers are increased, but activation is more important. Dig Dis Sci. 2014;59(5):897–898.Ref Type: Journal (Full). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Butterfield JH, Ravi A, Pongdee T. Mast cell mediators of significance in clinical practice in mastocytosis. Immunol Allergy Clin North Am. 2018;38:397–410. [DOI] [PubMed] [Google Scholar]
- 158.Samorapoompichit P, Kiener HP, Schernthaner GH, et al. Detection of tryptase in cytoplasmic granules of basophils in patients with chronic myeloid leukemia and other myeloid neoplasms. Blood. 2001;98:2580–2583. [DOI] [PubMed] [Google Scholar]
- 159.Jogie-Brahim S, Min HK, Fukuoka Y, et al. Expression of alpha-tryptase and beta-tryptase by human basophils. J Allergy Clin Immunol. 2004;113:1086–1092. [DOI] [PubMed] [Google Scholar]
- 160.Afrin LB, Molderings GJ. A concise, practical guide to diagnostic assessment for mast cell activation disease. World J Hematol. 2014;3:1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Shanmugam G, Schwartz LB, Khan DA. Prolonged elevation of serum tryptase in idiopathic anaphylaxis. J Allergy Clin Immunol. 2006;117:950–951. [DOI] [PubMed] [Google Scholar]
- 162.Schwartz LB. Diagnostic value of tryptase in anaphylaxis and mastocytosis. Immunol Allergy Clin North Am. 2006;26:451–463. [DOI] [PubMed] [Google Scholar]
- 163.Lin RY, Schwartz LB, Curry A, et al. Histamine and tryptase levels in patients with acute allergic reactions: an emergency department-based study. J Allergy Clin Immunol. 2000;106:65–71. [DOI] [PubMed] [Google Scholar]
- 164.Oosting E, Neugebauer E, Keyzer JJ, et al. Determination of histamine in human plasma: the European external quality control study 1988. Clin Exp Allergy. 1990;20:349–357. [DOI] [PubMed] [Google Scholar]
- 165.Awad JA, Morrow JD, Roberts LJ. Detection of the major urinary metabolite of prostaglandin D2 in the circulation: demonstration of elevated levels in patients with disorders of systemic mast cell activation. J Allergy Clin Immunol. 1994;93:817–824. [DOI] [PubMed] [Google Scholar]
- 166.Ravi A, Butterfield J, Weiler CR. Mast cell activation syndrome: improved identification by combined determinations of serum tryptase and 24-hour urine 11beta-prostaglandin2alpha. J Allergy Clin Immunol Pract. 2014;2:775–778. [DOI] [PubMed] [Google Scholar]
- 167.Vysniauskaite M, Hertfelder HJ, Oldenburg J, et al. Determination of plasma heparin level improves identification of systemic mast cell activation disease. PLoS ONE. 2015;10:e0124912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Theoharides TC, Boucher W, Spear K. Serum interleukin-6 reflects disease severity and osteoporosis in mastocytosis patients. Int Arch Allergy Immunol. 2002;128:344–350. [DOI] [PubMed] [Google Scholar]
- 169.Brockow K, Akin C, Huber M, et al. IL-6 levels predict disease variant and extent of organ involvement in patients with mastocytosis. Clin Immunol. 2005;115:216–223. [DOI] [PubMed] [Google Scholar]
- 170.Mayado A, Teodosio C, Garcia-Montero AC, et al. Increased IL6 plasma levels in indolent systemic mastocytosis patients are associated with high risk of disease progression. Leukemia. 2015;30:124–130. [DOI] [PubMed] [Google Scholar]
- 171.Greiner G, Witzeneder N, Berger A, et al. CCL2 is a KIT D816V-dependent modulator of the bone marrow microenvironment in systemic mastocytosis. Blood. 2017;129:371–382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Maintz L, Wardelmann E, Walgenbach K, et al. Neuropeptide blood levels correlate with mast cell load in patients with mastocytosis. Allergy. 2011;66:862–869. [DOI] [PubMed] [Google Scholar]
- 173.••.Levi-Schaffer F, Scheffel J. Dangerous liaisons in anaphylaxis. Science. 2018;362:640–641. Important discussion on the role of dendritic cells and exosomes in the pathogenesis of anaphylaxis. [DOI] [PubMed] [Google Scholar]
- 174.Skokos D, Botros HG, Demeure C, et al. Mast cell-derived exosomes induce phenotypic and functional maturation of dendritic cells and elicit specific immune responses in vivo. J Immunol. 2003;170:3037–3045. [DOI] [PubMed] [Google Scholar]
- 175.Bryniarski K, Ptak W, Jayakumar A, et al. Antigen-specific, antibody-coated, exosome-like nanovesicles deliver suppressor T-cell microRNA-150 to effector T cells to inhibit contact sensitivity. J Allergy Clin Immunol. 2013;132:170–181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Halova I, Ronnberg E, Draberova L, et al. Changing the threshold-Signals and mechanisms of mast cell priming. Immunol Rev. 2018;282:73–86. [DOI] [PubMed] [Google Scholar]
- 177.Alysandratos KD, Asadi S, Angelidou A, et al. Neurotensin and CRH interactions augment human mast cell activation. PloS One. 2012;7:e48934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Asadi S, Alysandratos KD, Angelidou A, et al. Substance P (SP) induces expression of functional corticotropin-releasing hormone receptor-1 (CRHR-1) in human mast cells. J Invest Dermatol. 2012;132:324–329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Drube S, Heink S, Walter S, et al. The receptor tyrosine kinase c-Kit controls IL-33 receptor signaling in mast cells. Blood. 2010;115:3899–3906. [DOI] [PubMed] [Google Scholar]
- 180.Kounis NG, Hahalis G, Theoharides TC. Coronary stents, hypersensitivity reactions and the Kounis syndrome. J Invasive Cardiol. 2007;20:314–323. [DOI] [PubMed] [Google Scholar]
- 181.Paratz ED, Khav N, Burns AT. Systemic mastocytosis, kounis syndrome and coronary intervention: case report and systematic review. Heart Lung Circ. 2017;26:772–778. [DOI] [PubMed] [Google Scholar]
- 182.Rossi S, Pitidis A. Multiple chemical sensitivity: review of the state of the art in epidemiology, diagnosis, and future perspectives. J Occup Environ Med. 2018;60:138–146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Theoharides TC, Tsilioni I, Patel AB, et al. Atopic diseases and inflammation of the brain in the pathogenesis of autism spectrum disorders. Transl Psychiatry. 2016;6:e844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.•.Theoharides TC. Autism spectrum disorders and mastocytosis. Int J Immunopathol Pharmacol. 2009;22:859–865. First report that children with mastocytosis have a higher risk of developing autism. [DOI] [PubMed] [Google Scholar]
- 185.Fombonne E Epidemiology of pervasive developmental disorders. Pediatr Res. 2009;65:591–598. [DOI] [PubMed] [Google Scholar]
- 186.McPartland J, Volkmar FR. Autism and related disorders. Handb Clin Neurol. 2012;106:407–418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Lai MC, Lombardo MV, Autism B-CS. Lancet. 2014;383:896–910. [DOI] [PubMed] [Google Scholar]
- 188.Mostafa GA, Hamza RT, El-Shahawi HH. Allergic manifestations in autistic children: relation to disease severity. J Pediatr Neurol. 2008;6:115–123. [Google Scholar]
- 189.Otani IM, Carroll RW, Yager P, et al. Diffuse cutaneous mastocytosis with novel somatic KIT mutation K509I and association with tuberous sclerosis. Clin Case Rep. 2018;6:1834–1840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Kotey S, Ertel K, Whitcomb B. Co-occurrence of autism and asthma in a nationally-representative sample of children in the United States. J Autism Dev Disord. 2014;44:3083–3088. [DOI] [PubMed] [Google Scholar]
- 191.Lyall K, de WJ V, Ashwood P, et al. Allergies in children with autism spectrum disorders: results from the CHARGE study. Autism Res. 2015;8:567–574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Zerbo O, Leong A, Barcellos L, et al. Immune mediated conditions in autism spectrum disorders. Brain Behav Immun. 2015;46:232–236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Chen SW, Zhong XS, Jiang LN, et al. Maternal autoimmune diseases and the risk of autism spectrum disorders in offspring: a systematic review and meta-analysis. Behav Brain Res. 2015;296:61–69. [DOI] [PubMed] [Google Scholar]
- 194.Billeci L, Tonacci A, Tartarisco G, et al. Association between atopic dermatitis and autism spectrum disorders: a systematic review. Am J Clin Dermatol. 2015;16:371–388. [DOI] [PubMed] [Google Scholar]
- 195.Wills S, Cabanlit M, Bennett J, et al. Detection of autoantibodies to neural cells of the cerebellum in the plasma of subjects with autism spectrum disorders. Brain Behav Immun. 2008;23:64–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Rossi CC, de WJ V, Rogers SJ, et al. Detection of plasma autoantibodies to brain tissue in young children with and without autism spectrum disorders. Brain Behav Immun. 2011;25:1123–1135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Mostafa GA, Al-Ayadhi LY. The possible relationship between allergic manifestations and elevated serum levels of brain specific auto-antibodies in autistic children. J Neuroimmunol. 2013;261:77–81. [DOI] [PubMed] [Google Scholar]
- 198.Netea MG, Balkwill F, Chonchol M, et al. A guiding map for inflammation. Nat Immunol. 2017;18:826–831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Rider P, Voronov E, Dinarello CA, et al. Alarmins: feel the stress. J Immunol. 2017;198:1395–1402. [DOI] [PubMed] [Google Scholar]
- 200.Ransohoff RM, Brown MA. Innate immunity in the central nervous system. J Clin Invest. 2012;122:1164–1171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Skaper SD, Giusti P, Facci L. Microglia and mast cells: two tracks on the road to neuroinflammation. FASEB J. 2012;26:3103–3117. [DOI] [PubMed] [Google Scholar]
- 202.Forsythe P Mast cells in neuroimmune interactions. Trends Neurosci. 2018. [DOI] [PubMed] [Google Scholar]
- 203.Zhang X, Wang Y, Dong H, et al. Induction of microglial activation by mediators released from mast cells. Cell Physiol Biochem. 2016;38:1520–1531. [DOI] [PubMed] [Google Scholar]
- 204.Estes ML, McAllister AK. Immune mediators in the brain and peripheral tissues in autism spectrum disorder. Nat Rev Neurosci. 2015;16:469–486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Theoharides TC, Stewart JM, Panagiotidou S, et al. Mast cells, brain inflammation and autism. Eur J Pharmacol. 2015;778:96–102. [DOI] [PubMed] [Google Scholar]
- 206.Edvinsson L, Cervos-Navarro J, Larsson LI, et al. Regional distribution of mast cells containing histamine, dopamine or 5-hydroxytryptamine in the mammalian brain. Neurology. 1977;27:878–884. [DOI] [PubMed] [Google Scholar]
- 207.Pang X, Letourneau R, Rozniecki JJ, et al. Definitive characterization of rat hypothalamic mast cells. Neuroscience. 1996;73(3):889–902. [DOI] [PubMed] [Google Scholar]
- 208.Theoharides TC. Mast cells: the immune gate to the brain. Life Sci. 1990;46:607–617. [DOI] [PubMed] [Google Scholar]
- 209.Theoharides TC, Konstantinidou A. Corticotropin-releasing hormone and the blood-brain-barrier. Front Biosci. 2007;12:1615–1628. [DOI] [PubMed] [Google Scholar]
- 210.Esposito P, Chandler N, Kandere-Grzybowska K, et al. Corticotropin-releasing hormone (CRH) and brain mast cells regulate blood-brain-barrier permeability induced by acute stress. J Pharmacol Exp Ther. 2002;303:1061–1066. [DOI] [PubMed] [Google Scholar]
- 211.Ribatti D The crucial role of mast cells in blood-brain barrier alterations. Exp Cell Res. 2015;338:119–125. [DOI] [PubMed] [Google Scholar]
- 212.Polyzoidis S, Koletsa T, Panagiotidou S, et al. Mast cells in meningiomas and brain inflammation. J Neuroinflammation. 2015;12:170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Molderings GJ, Haenisch B, Bogdanow M, et al. Familial occurrence of systemic mast cell activation disease. PloS One. 2013;8:e76241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Scherber RM, Borate U. How we diagnose and treat systemic mastocytosis in adults. Br J Haematol. 2018;180:11–23. [DOI] [PubMed] [Google Scholar]
- 215.Siebenhaar F, Redegeld FA, Bischoff SC, et al. Mast Cells as Drivers of Disease and Therapeutic Targets. Trends Immunol. 2018;39(2):151–162. [DOI] [PubMed] [Google Scholar]
- 216.Molderings GJ, Haenisch B, Brettner S, et al. Pharmacological treatment options for mast cell activation disease. Naunyn Schmiedebergs Arch Pharmacol. 2016;389:671–694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Alvarez-Twose I, Gonzalez-de-Olano D, Sanchez-Munoz L, et al. Validation of the REMA score for predicting mast cell clonality and systemic mastocytosis in patients with systemic mast cell activation symptoms. Int Arch Allergy Immunol. 2012;157:275–280. [DOI] [PubMed] [Google Scholar]
- 218.Kettelhut BV, Berkebile C, Bradley D, et al. A double-blind, placebo-controlled, crossover trial of ketotifen versus hydroxyzine in the treatment of pediatric mastocytosis. J Allergy Clin Immunol. 1989;83:866–870. [DOI] [PubMed] [Google Scholar]
- 219.Schoch C In vitro inhibition of human conjunctival mast-cell degranulation by ketotifen. J Ocul Pharmacol Ther. 2003;19:75–81. [DOI] [PubMed] [Google Scholar]
- 220.Vasiadi M, Kalogeromitros K, Kempuraj D, et al. Rupatadine inhibits pro-inflammatory mediator secretion from human mast cells triggered by different stimuli. Int Arch Allergy Immunol. 2010;151:38–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Alevizos M, Karagkouni A, Vasiadi M, et al. Rupatadine inhibits inflammatory mediator release from human LAD2 cultured mast cells stimulated by PAF. Ann Allergy Asthma Immunol. 2013;111:524–527. [DOI] [PubMed] [Google Scholar]
- 222.Wolff SC, Brubaker KNTFEHLJRBJ. Anticholinergic effects of antihistamine drugs used in the clinic. J Allergy Clin Immunol. 2007;119. [Google Scholar]
- 223.Shimizu T, Tokuda Y. Acute urinary retention. Intern Med. 2011;50:2255. [DOI] [PubMed] [Google Scholar]
- 224.Gray SL, Anderson ML, Dublin S, et al. Cumulative use of strong anticholinergics and incident dementia: a prospective cohort study. JAMA Intern Med. 2015;175:401–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Theoharides TC, Stewart JM. Antihistamines and mental status. J Clin Psychopharmacol. 2016;36:195–197. [DOI] [PubMed] [Google Scholar]
- 226.Zhang T, Finn DF, Barlow JW, et al. Mast cell stabilisers. Eur J Pharmacol. 2016;778:158–168. [DOI] [PubMed] [Google Scholar]
- 227.Theoharides TC, Sieghart W, Greengard P, et al. Antiallergic drug cromolyn may inhibit histamine secretion by regulating phosphorylation of a mast cell protein. Science. 1980;207:80–82. [DOI] [PubMed] [Google Scholar]
- 228.Oka T, Kalesnikoff J, Starkl P, et al. Evidence questioning cromolyn’s effectiveness and selectivity as a ‘mast cell stabilizer’ in mice. Lab Invest. 2012;92:1472–1482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Vieira Dos SR, Magerl M, Martus P, et al. Topical sodium cromoglicate relieves allergen- and histamine-induced dermal pruritus. Br J Dermatol. 2010;162:674–676. [DOI] [PubMed] [Google Scholar]
- 230.Butterfield JH, Weiler CR. Prevention of mast cell activation disorder-associated clinical sequelae of excessive prostaglandin D(2) production. Int Arch Allergy Immunol. 2008;147:338–343. [DOI] [PubMed] [Google Scholar]
- 231.Conti P, Varvara G, Murmura G, et al. Comparison of beneficial actions of non-steroidal anti-inflammatory drugs to flavonoids. J Biol Regul Homeost Agents. 2013;27:1–7. [PubMed] [Google Scholar]
- 232.Varvara G, Tettamanti L, Gallenga CE, et al. Stimulated mast cells release inflammatory cytokines: potential suppression and therapeutical aspects. J Biol Regul Homeost Agents. 2018;32:1355–1360. [PubMed] [Google Scholar]
- 233.Butterfield JH, Kao PC, Klee GC, et al. Aspirin idiosyncrasy in systemic mast cell disease: a new look at mediator release during aspirin desensitization. Mayo Clin Proc. 1995;70:481–487. [DOI] [PubMed] [Google Scholar]
- 234.Molderings GJ, Raithel M, Kratz F, et al. Omalizumab treatment of systemic mast cell activation disease: experiences from four cases. Intern Med. 2011;50:611–615. [DOI] [PubMed] [Google Scholar]
- 235.Jayapal M, Tay HK, Reghunathan R, et al. Genome-wide gene expression profiling of human mast cells stimulated by IgE or FcepsilonRI-aggregation reveals a complex network of genes involved in inflammatory responses. BMC Genomics. 2006;7:210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Kalogeromitros D, Makris M, Koti I, et al. A simple 3-day “rush” venom immunotherapy protocol: documentation of safety. Allergol Immunopathol (Madr). 2010;38:69–73. [DOI] [PubMed] [Google Scholar]
- 237.Gotlib J, Kluin-Nelemans HC, George TI, et al. Safety of midostaurin in advanced systemic mastocytosis. N Engl J Med. 2016;374:2530–2541. [DOI] [PubMed] [Google Scholar]
- 238.Rapid responses to avapritinib (BLU-285) in mastocytosis. Cancer Discov. 2018;8:133. [DOI] [PubMed] [Google Scholar]
- 239.Guo W, Kong E, Meydani M. Dietary polyphenols, inflammation, and cancer. Nutr Cancer. 2009;61:807–810. [DOI] [PubMed] [Google Scholar]
- 240.Middleton EJ, Kandaswami C, Theoharides TC. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease and cancer. Pharmacol Rev. 2000;52:673–751. [PubMed] [Google Scholar]
- 241.Xiao ZP, Peng ZY, Peng MJ, et al. Flavonoids health benefits and their molecular mechanism. Mini Rev Med Chem. 2011;11:169–177. [DOI] [PubMed] [Google Scholar]
- 242.Taliou A, Zintzaras E, Lykouras L, et al. An open-label pilot study of a formulation containing the anti-inflammatory flavonoid luteolin and its effects on behavior in children with autism spectrum disorders. Clin Ther. 2013;35:592–602. [DOI] [PubMed] [Google Scholar]
- 243.Kimata M, Shichijo M, Miura T, et al. Effects of luteolin, quercetin and baicalein on immunoglobulin E-mediated mediator release from human cultured mast cells. Clin Exp Allergy. 2000;30:501–508. [DOI] [PubMed] [Google Scholar]
- 244.Asadi S, Zhang B, Weng Z, et al. Luteolin and thiosalicylate inhibit HgCl(2) and thimerosal-induced VEGF release from human mast cells. Int J Immunopathol Pharmacol. 2010;23:1015–1020. [DOI] [PubMed] [Google Scholar]
- 245.Weng Z, Patel AB, Panagiotidou S, et al. The novel flavone tetramethoxyluteolin is a potent inhibitor of human mast cells. J Allergy Clin Immunol. 2015;135:1044–1052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Lee JY, Kim JM, Kim CJ. Flavones derived from nature attenuate the immediate and late-phase asthmatic responses to aerosolized-ovalbumin exposure in conscious guinea pigs. Inflamm Res. 2014;63:53–60. [DOI] [PubMed] [Google Scholar]
- 247.Jin M, Son KH, Chang HW. Luteolin-7-O-glucoside suppresses leukotriene C(4) production and degranulation by inhibiting the phosphorylation of mitogen activated protein kinases and phospholipase Cgamma1 in activated mouse bone marrow-derived mast cells. Biol Pharm Bull. 2011;34:1032–1036. [DOI] [PubMed] [Google Scholar]
- 248.Patel AB, Tsilioni I, Weng Z, et al. TNF stimulates IL-6, CXCL8 and VEGF secretion from human keratinocytes via activation of mTOR, inhibited by tetramethoxyluteolin. Exp Dermatol. 2018;27:135–143. [DOI] [PubMed] [Google Scholar]
- 249.Sharma V, Mishra M, Ghosh S, et al. Modulation of interleukin-1beta mediated inflammatory response in human astrocytes by flavonoids: implications in neuroprotection. Brain Res Bull. 2007;73:55–63. [DOI] [PubMed] [Google Scholar]
- 250.Jang S, Kelley KW, Johnson RW. Luteolin reduces IL-6 production in microglia by inhibiting JNK phosphorylation and activation of AP-1. Proc Natl Acad Sci USA. 2008;105:7534–7539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Patel AB, Tsilioni I, Leeman SE, et al. Neurotensin stimulates sortilin and mTOR in human microglia inhibitable by methoxyluteolin, a potential therapeutic target for autism. Proc Natl Acad Sci U S A. 2016;113:E7049–E7058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Dirscherl K, Karlstetter M, Ebert S, et al. Luteolin triggers global changes in the microglial transcriptome leading to a unique anti-inflammatory and neuroprotective phenotype. J Neuroinflammation. 2010;7:3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Jang S, Dilger RN, Johnson RW. Luteolin inhibits microglia and alters hippocampal-dependent spatial working memory in aged mice. J Nutr. 2010;140:1892–1898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Kao TK, Ou YC, Lin SY, et al. Luteolin inhibits cytokine expression in endotoxin/cytokine-stimulated microglia. J Nutr Biochem. 2011;22:612–624. [DOI] [PubMed] [Google Scholar]
- 255.Theoharides TC, Asadi S, Panagiotidou S. A case series of a luteolin formulation (Neuroprotek®) in children with autism spectrum disorders. Int J Immunopathol Pharmacol. 2012;25:317–323. [DOI] [PubMed] [Google Scholar]
- 256.Seelinger G, Merfort I, Schempp CM. Anti-oxidant, anti-inflammatory and anti-allergic activities of luteolin. Planta Med. 2008;74:1667–1677. [DOI] [PubMed] [Google Scholar]
- 257.Nabavi SF, Braidy N, Gortzi O, et al. Luteolin as an anti-inflammatory and neuroprotective agent: a brief review. Brain Res Bull. 2015;119:1–11. [DOI] [PubMed] [Google Scholar]
- 258.Chen HQ, Jin ZY, Wang XJ, et al. Luteolin protects dopaminergic neurons from inflammation-induced injury through inhibition of microglial activation. Neurosci Lett. 2008;448:175–179. [DOI] [PubMed] [Google Scholar]
- 259.Franco JL, Posser T, Missau F, et al. Structure-activity relationship of flavonoids derived from medicinal plants in preventing methylmercury-induced mitochondrial dysfunction. Environ Toxicol Pharmacol. 2010;30:272–278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Xu SL, Bi CW, Choi RC, et al. Flavonoids induce the synthesis and secretion of neurotrophic factors in cultured rat astrocytes: a signaling response mediated by estrogen receptor. Evid Based Complement Alternat Med. 2013;2013:127075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Jang SW, Liu X, Yepes M, et al. A selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavone. Proc Natl Acad Sci USA. 2010;107:2687–2692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Johnson RA, Lam M, Punzo AM, et al. 7,8-dihydroxyflavone exhibits therapeutic efficacy in a mouse model of Rett syndrome. J Appl Physiol. 2012;112:704–710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Fatokun AA, Liu JO, Dawson VL, et al. Identification through high-throughput screening of 4ʹ-methoxyflavone and 3ʹ,4ʹ-dimethoxyflavone as novel neuroprotective inhibitors of parthanatos. Br J Pharmacol. 2013;169:1263–1278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Kang OH, Choi JG, Lee JH, et al. Luteolin isolated from the flowers of Lonicera japonica suppresses inflammatory mediator release by blocking NF-kappaB and MAPKs activation pathways in HMC-1 cells. Molecules. 2010;15:385–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Yang Y, Oh JM, Heo P, et al. Polyphenols differentially inhibit degranulation of distinct subsets of vesicles in mast cells by specific interaction with granule-type-dependent SNARE complexes. Biochem J. 2013;450:537–546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.Theoharides TC. Luteolin as a therapeutic option for multiple sclerosis. J Neuroinflammation. 2009;6:29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267.Theoharides TC, Tsilioni I. Tetramethoxyluteolin for the treatment of neurodegenerative diseases. Curr Top Med Chem. 2018. [DOI] [PubMed] [Google Scholar]
- 268.Ashaari Z, Hadjzadeh MA, Hassanzadeh G, et al. The flavone luteolin improves central nervous system disorders by different mechanisms: a review. J Mol Neurosci. 2018;65:491–506. [DOI] [PubMed] [Google Scholar]
- 269.Prasain JK, Barnes S. Metabolism and bioavailability of flavonoids in chemoprevention: current analytical strategies and future prospectus. Mol Pharm. 2007;4:846–864. [DOI] [PubMed] [Google Scholar]
- 270.Walle T Absorption and metabolism of flavonoids. Free Radic Biol Med. 2004;36:829–837. [DOI] [PubMed] [Google Scholar]
- 271.Graefe EU, Derendorf H, Veit M. Pharmacokinetics and bioavailability of the flavonol quercetin in humans. Int J Clin Pharmacol Ther. 1999;37:219–233. [PubMed] [Google Scholar]
- 272.Chen Z, Zheng S, Li L, et al. Metabolism of flavonoids in human: a comprehensive review. Curr Drug Metab. 2014;15:48–61. [DOI] [PubMed] [Google Scholar]
- 273.Walle T Methylation of dietary flavones greatly improves their hepatic metabolic stability and intestinal absorption. Mol Pharm. 2007;4:826–832. [DOI] [PubMed] [Google Scholar]
- 274.Corcoran MP, McKay DL, Blumberg JB. Flavonoid basics: chemistry, sources, mechanisms of action, and safety. J Nutr Gerontol Geriatr. 2012;31:176–189. [DOI] [PubMed] [Google Scholar]
- 275.Harwood M, Nielewska-Nikiel B, Borzelleca JF, et al. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem Toxicol. 2007;45:2179–2205. [DOI] [PubMed] [Google Scholar]
- 276.Kawanishi S, Oikawa S, Murata M. Evaluation for safety of antioxidant chemopreventive agents. Antioxid Redox Signal. 2005;7:1728–1739. [DOI] [PubMed] [Google Scholar]
- 277.Theoharides TC, Conti P, Economu M. Brain inflammation, neuropsychiatric disorders, and immunoendocrine effects of luteolin. J Clin Psychopharmacol. 2014;34:187–189. [DOI] [PubMed] [Google Scholar]
- 278.Domitrovic R, Jakovac H, Milin C, et al. Dose- and time-dependent effects of luteolin on carbon tetrachloride-induced hepatotoxicity in mice. Exp Toxicol Pathol. 2009;61:581–589. [DOI] [PubMed] [Google Scholar]
- 279.Weng Z, Zhang B, Asadi S, et al. Quercetin is more effective than cromolyn in blocking human mast cell cytokine release and inhibits contact dermatitis and photosensitivity in humans. PloS One. 2012;7:e33805-k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Tsilioni I, Taliou A, Francis K, et al. Children with autism spectrum disorders, who improved with a luteolin containing dietary formulation, show reduced serum levels of TNF and IL-6. Transl Psychiatry. 2015;5:e647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 281.Theoharides TC, Stewart JM. Post-lyme syndrome-associated polyneuropathy treated with immune immunoglobulin and a luteolin-containing formulation. J Clin Psychopharmacol. 2016;36:290–291. [DOI] [PubMed] [Google Scholar]
- 282.Theoharides TC, Stewart JM, Tsilioni I. Tolerability and benefit of a tetramethoxyluteolin-containing skin lotion. Int J Immunopathol Pharmacol. 2017;30:146–151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Theoharides TC. Vitamin D and Atopy. Clin Ther. 2017;39:880–883. [DOI] [PubMed] [Google Scholar]
- 284.Clemons A, Vasiadi M, Kempuraj D, et al. Amitriptyline and prochlorperazine inhibit proinflammatory mediator release from human mast cells: possible relevance to chronic fatigue syndrom. J Clin Psychopharmacol. 2011;31:385–387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Krystel-Whittemore M, Dileepan KN, Wood JG. Mast cell: a multi-functional master cell. Front Immunol. 2015;6:620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Olivera A, Beaven MA, Metcalfe DD. Mast cells signal their importance in health and disease. J Allergy Clin Immunol. 2018;142:381–393. [DOI] [PubMed] [Google Scholar]
- 287.Theoharides TC, Leeman SE. Effect of IL-33 on de novo synthesized mediators from human mast cells. J Allergy Clin Immunol. 2018. [DOI] [PubMed] [Google Scholar]
- 288.Caslin HL, Kiwanuka KN, Haque TT, et al. Controlling mast cell activation and homeostasis: work influenced by bill paul that continues today. Front Immunol. 2018;9:868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Seok J, Warren HS, Cuenca AG, et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Pnas. 2013;110:3507–3512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290.Frossi B, Mion F, Sibilano R, et al. Is it time for a new classification of mast cells? What do we know about mast cell heterogeneity? Immunol Rev. 2018;282:35–46. [DOI] [PubMed] [Google Scholar]
- 291.Tellechea A, Leal EC, Kafanas A, et al. Mast cells regulate wound healing in diabetes. Diabetes. 2016;65:2006–2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Ribatti D Mast cells as therapeutic target in cancer. Eur J Pharmacol. 2015. [DOI] [PubMed] [Google Scholar]
- 293.Silver R, Ramos CL, Silverman AJ. Sexual behavior triggers the appearance of non-neuronal cells containing gonadotropin-releasing hormone-like immunoreactivity. J Neuroendocrinol. 1991;4:1–3. [DOI] [PubMed] [Google Scholar]
- 294.••.Lenz KM, Pickett LA, Wright CL, et al. Mast cells in the developing brain determine adult sexual behavior. J Neurosci. 2018;38:8044–8059. This paper demonstrates that mast cells may define adult sexual behavior at least in in rodents. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Kim DK, Cho YE, Komarow HD, et al. Mastocytosis-derived extracellular vesicles exhibit a mast cell signature, transfer KIT to stellate cells, and promote their activation. Proc Natl Acad Sci U S A. 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Haenisch B, Bonisch H, Cichon S, et al. Effects of exogenous agmatine in human leukemia HMC-1 and HL-60 cells on proliferation, polyamine metabolism and cell cycle. Leuk Res. 2011;35:1248–1253. [DOI] [PubMed] [Google Scholar]
- 297.Vliagoftis H, Boucher WS, Mak LL, et al. Inhibition of mast cell secretion by oxidation products of natural polyamines. Biochem Pharmacol. 1992;43:2237–2245. [DOI] [PubMed] [Google Scholar]
- 298.Garcia-Faroldi G, Rodriguez CE, Urdiales JL, et al. Polyamines are present in mast cell secretory granules and are important for granule homeostasis. PLoS ONE. 2010;5:e15071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299.Gross AR, Theoharides TC. Chondroitin sulfate inhibits secretion of TNF and CXCL8 from human mast cells stimulated by IL-33. Biofactors. 2018;1–13. [DOI] [PubMed] [Google Scholar]
- 300.Abramson J, Licht A, Pecht I. Selective inhibition of the Fc epsilon RI-induced de novo synthesis of mediators by an inhibitory receptor. Embo J. 2006;25:323–334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Dinarello CA, Bufler P. Interleukin-37. Semin Immunol. 2013;25:466–468. [DOI] [PubMed] [Google Scholar]
- 302.Dinarello CA, Nold-Petry C, Nold M, et al. Suppression of innate inflammation and immunity by interleukin-37. Eur J Immunol. 2016;46:1067–1081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 303.Tettamanti L, Kritas SK, Gallenga CE, et al. IL-33 mediates allergy through mast cell activation: potential inhibitory effect of certain cytokines. J Biol Regul Homeost Agents. 2018;32:1061–1065. [PubMed] [Google Scholar]
- 304.Mastrangelo F, Frydas I, Ronconi G, et al. Low-grade chronic inflammation mediated by mast cells in fibromyalgia: role of IL-37. J Biol Regul Homeost Agents. 2018;32:195–198. [PubMed] [Google Scholar]