

Abstract
Background
Common bacterial blight (CBB) caused by Xanthomonas phaseoli pv. phaseoli and Xanthomonas citri pv. fuscans is one of the major threats to common bean crops (Phaseolus vulgaris L.). Resistance to CBB is particularly complex as 26 quantitative resistance loci to CBB have been described so far. To date, transcriptomic studies after CBB infection have been very scarce and the molecular mechanisms underlying susceptibility or resistance are largely unknown.
Results
We sequenced and annotated the genomes of two common bean genotypes being either resistant (BAT93) or susceptible (JaloEEP558) to CBB. Reciprocal BLASTp analysis led to a list of 20,787 homologs between these genotypes and the common bean reference genome (G19833), which provides a solid dataset for further comparative analyses. RNA-Seq after inoculation with X. phaseoli pv. phaseoli showed that the susceptible genotype initiated a more intense and diverse biological response than the resistant genotype. Resistance was linked to upregulation of the salicylic acid pathway and downregulation of photosynthesis and sugar metabolism, while susceptibility was linked to downregulation of resistance genes and upregulation of the ethylene pathway and of genes involved in cell wall modification.
Conclusions
This study helps better understanding the mechanisms occurring during the early colonization phase of common bean by Xanthomonas and unveils new actors potentially important for resistance and susceptibility to CBB. We discuss the potential link between the pathways induced during bean colonization and genes induced by transcription activator-like effectors (TALEs), as illustrated in other Xanthomonas pathovars.
Keywords: Common bean, Xanthomonas, Common bacterial blight, RNA-Seq, Resistance
Background
Plant immunity is governed by a two-tier system capable of monitoring the presence of pathogens [1, 2]. The first layer of the plant immune system consists of the recognition of evolutionarily conserved pathogen- or microbe-associated molecular patterns (PAMPs or MAMPs) by pattern-recognition receptors (PRRs) [3, 4]. PRRs belong typically to the receptor-like kinase (RLK) family, which encompasses membrane proteins with an extracellular domain composed of either leucine-rich repeats or lysin motifs [5]. Activation of PRRs leads to a complex response called PAMP-triggered immunity (PTI) involving intracellular signaling, transcriptional reprogramming, and biosynthesis of compounds that limit microbial colonization [6]. Pathogens employ an array of proteins called effectors, a large proportion of which are able to subvert PTI [7]. For example, bacterial pathogens use a type III secretion system to inject dozens of type III effectors into host cells [8–10]. The second layer of plant immunity or effector-triggered immunity (ETI) consists of the direct or indirect recognition of effectors by intracellular disease resistance (R) proteins [11]. Most of these proteins belong to a large family of nucleotide-binding leucine-rich repeat (NLR) receptors. After pathogen detection, plants usually trigger a burst of Ca2+ followed by an extracellular production of reactive oxygen species and activation of mitogen-activated protein kinases (MAPKs). Different molecular pathways can be activated, involving major hormones such as salicylic acid (SA), jasmonic acid (JA) and ethylene, which are key for regulating the immune response to pathogen [12–14]. These pathways lead to defense responses characterized by, but not limited to the production of pathogenesis-related (PR) proteins and cell wall reinforcement [15].
Common bean (Phaseolus vulgaris L.) is the main grain legume used for direct human consumption. This crop is of particular interest for human nutrition, as it is one of the main sources of protein in many countries from America and Africa, as well as a source of vitamins, fibers and minerals [16, 17]. Common bean originated from Mesoamerica, from where populations migrated to the Andean region 146,000 to 184,000 years ago, resulting in the creation of two major gene pools (Mesoamerican and Andean) [18]. Wild plants from both gene pools differ in morphology, Mesoamerican having thinner leaves and producing pods and seeds that are smaller, though more numerous than their Andean counterpart [19].
Common bacterial blight of bean (CBB) is one of the main threats to bean cultivation with yield losses of more than 40% under favourable conditions [20, 21] and up to 75% in the most severe cases [22, 23]. CBB is characterized by water-soaking spots on leaves, stems and pods, further evolving into necrotic lesions sometimes surrounded by a chlorotic halo in leaves. This disease is caused by Xanthomonas phaseoli pv. phaseoli and Xanthomonas citri pv. fuscans [24–27]. These bacteria are mainly transmitted by seeds, and occur in all regions where common bean is cultivated [22]. CBB is mainly controlled by prophylactic methods such as the use of pathogen-free seeds, two-year rotations with non-leguminous crops and burning of plant residues [28]. In quarantine areas, seed lots are routinely tested using a method involving isolation of bacterial strains and identification using specific PCR or pathogenicity tests [29, 30]. However, X. citri pv. fuscans and X. phaseoli pv. phaseoli were recently removed from the quarantine list of the European Union (EU delegated regulation 2019/2072), which was previously the main quarantine zone for CBB agents worldwide. In this context, it is important to develop and use resistant genotypes, which is the most economic and ecologically safe management strategy against CBB [31, 32].
Although variations of CBB resistance levels have been observed in several common bean accessions, no major R gene to CBB has been described so far [33–35]. On the other hand, several R gene to CBB have been described in P. acutifolius and P. coccineus [36, 37]. Introgression of CBB resistance from P. acutifolius in XAN lines led to two major quantitative resistance loci (QRLs) associated with markers SU91 and BC420 [38, 39], which explain a significant part of the phenotypic variation [40–42]. Most resistant cultivars bred for CBB resistance possess the SAP6 marker deriving from the Great Northern landrace cultivars, such as ‘Montana No.5’ and ‘GN #1 sel 27’ [43, 44]. So far, 26 QRLs to CBB have been mapped using nine different bi-parental populations. These QRLs are dispersed throughout the common bean genome and poorly co-localize with each other when comparing the different populations tested. Moreover, most CBB resistances vary according to plant maturity, plant organs (leaf, pod or seed), pathogenic strains and environment [34, 45, 46], which reflects the high complexity of CBB resistance.
Analyzing the transcriptomic response of common bean to CBB is a way to enhance our knowledge on the molecular mechanisms underlying CBB resistance, and can provide important information for developing genetic management of the disease. However, to our knowledge, transcriptomic studies during X. phaseoli pv. phaseoli infection are limited to a cDNA-AFLP analysis [47] and RT-qPCR assays focused on R genes [48, 49]. Previous work using X. phaseoli pv. phaseoli isolate W18 identified four QRLs to CBB in a cross between the Mesoamerican genotype BAT93 (resistant to CBB) and the Andean genotype JaloEEP558 (susceptible to CBB) [50]. These QRLs are located on chromosomes 2, 5, 7 and 9 and explain 75% of the phenotypic variation. Here, we studied the transcriptomic response of BAT93 and JaloEEP558 to X. phaseoli pv. phaseoli strain CFBP6546R 48 h after inoculation.
Results
Pathogenicity assays
Resistance and susceptible phenotypes of common bean genotypes BAT93 and JaloEEP558 were demonstrated with bacterial growth and symptom development after inoculation with X. phaseoli pv. phaseoli strain CFBP6546R. For both genotypes, bacterial population sizes decreased during the first day, then increased rapidly up to day 5 and stabilized over time, which is a typical dynamic for common bean colonization by Xanthomonas [51]. However, bacterial population sizes were significantly higher on JaloEEP558 than on BAT93 (p-value < 0.05) at 8 and 15 days post inoculation (Fig. 1A). Moreover, BAT93 presented almost no symptoms, while 39 to 50% of the leaf area were symptomatic on JaloEEP558 (Fig. 1B). Symptoms 15 days post inoculation (DPI) were characterized by the apparition of necrotic areas that were not detected at 8 DPI. These results show that JaloEEP558 is susceptible, while BAT93 is resistant to strain CFBP6546R, as previously described for X. phaseoli pv. phaseoli isolate W18 [50].
Fig. 1.

Pathogenicity of strain CFBP6546R on BAT93 and JaloEEP558. Bacterial population sizes over time on BAT93 (dotted line) and JaloEEP558 (full line) (a). Quantification of symptoms assessed by chlorophyll fluorescence imaging at 8 and 15 DPI (b). Total symptomatic areas corresponded to the sum of impacted, wilted and necrotic tissues defined by using Fv/Fm tresholds as previously compared to visual inspection [52]. Error bars represent the standard errors of the means for three biological replicates. Below the histogram are examples of the leaflets presenting symptoms representative of each condition, obtained by chlorophyll fluorescence imaging (top) or visible imaging (bottom). Letters indicate significantly different groups (Mann-Whitney test, p-value < 0.05). CFU: colony-forming units. gFM: grams of fresh materials. DPI: days post inoculation
Whole genome sequencing and annotation
The genomes of BAT93 and JaloEEP558 were sequenced and annotated to serve as basis for the mapping of corresponding RNA-Seq reads. For each genotype, Illumina sequencing produced around 3.2 × 108 paired reads totalizing 6.4 × 104 Mbp. The resulting assembly consisted of 36,622 scaffolds totalizing 453.0 Mbp for BAT93, and 31,483 scaffolds totalizing 449.3 Mbp for JaloEEP558, with a coverage of approximately 100X for both genotypes (Table 1). These assemblies represented around 84% of the P. vulgaris v2.1 reference genome (537.2 Mbp) from the Andean genotype G19833 [18] or 77% of the estimated genome size (~ 587 Mbp), and 82% of the BAT93 reference genome (549,6 Mbp) published by Vlasova et al. in 2016 [53]. Conversely, structural annotation predicted 33,275 and 32,914 protein coding genes for BAT93 and JaloEEP558, respectively, which are numbers higher than those found in the reference genomes of G19833 (27,433) or BAT93 (30,491). Reciprocal BLASTp (e-value ≤1 × 10− 6) using all predicted genes showed that our BAT93 and JaloEEP558 genome sequences shared more homologs (27,208) together than with both G19833 and BAT93 reference genomes (Table S1). Thus, to avoid possible biases due to different sequencing and annotation methods, we used our own version of the BAT93 genome for RNA-Seq mapping and comparative transcriptomic analysis between BAT93 and JaloEEP558. To ensure the quality of our data, we used a subset of homologs between BAT93 and JaloEEP558 that had the same best hit after BLASTp on the G19833 reference genome. After removal of paralogs, we obtained 20,787 unique genes that we used for all the analyses presented hereafter (Table S2).
Table 1.
Summary of sequencing, assembly and annotation data
| BAT93 | JaloEEP558 | |
|---|---|---|
| Whole genome sequencing | ||
| Assembly length (bp) | 452,993,439 | 449,275,055 |
| Coverage | 108x | 114x |
| Number of scaffolds | 36,622 | 31,483 |
| N50 (size/number) | 35,794/3086 | 44,310/2481 |
| N90 (size/number) | 4951/16,296 | 5936/13,165 |
| % of Ns | 0.92% | 0.84% |
| Annotation | ||
| Predicted genes | 33,275 | 32,914 |
| RNA sequencing | ||
| Average raw reads | 23,237,717 | 23,040,441 |
| Average mapped reads on genes | 20,197,275 | 20,731,141 |
| % of mapped reads on genes | 87% | 90% |
Analysis of differentially-expressed genes
To explore the responses of BAT93 and JaloEEP558 to strain CFBP6546R, transcriptomes of inoculated leaves were produced. In other Xanthomonas-plant pathosystems such as tomato [54], rice [55] or sweet orange [56, 57], bacterial effectors impacted plant transcriptomes around 24 to 48 h after leaf infiltration. Here, we performed RNA-Seq analyses on inoculated leaves 48 h after infiltration. Illumina sequencing led to a total of 139.4 and 138.2 million raw reads for BAT93 and JaloEEP558 respectively, with an average of 23.2 million raw reads per sample for BAT93 and 23.0 million raw reads per sample for JaloEEP558 (Table 1). After stringent quality check, data cleaning and mapping, we obtained an average of 20.2 million mapped reads on genes per sample (87% of raw reads) for BAT93 and 20.7 million mapped reads on genes per sample (90% of raw reads) for JaloEEP558. Water-inoculated and bacteria-inoculated plants formed distinct groups after principal component analysis, confirming the similarity of biological replicates within each condition and the similarity of both genotypes after water treatment (Fig. S1). Similar trends were observed using a Pearson correlation matrix, indicating that the bacteria had a significant effect on the transcriptomes of both genotypes. After bacterial inoculation, a total of 5581 out of the 20,787 homologs were differentially expressed in at least one genotype compared to water inoculation (Table S3), using adjusted p-value < 0.05 and |log2FC| > 1.5 (see Materials and Methods). Comparison of RT-qPCR and RNA-Seq values on 10 genes presenting different patterns of expression in BAT93 and JaloEEP558 revealed a high correlation for these genes (Pearson r = 0.95), further confirming the reliability of RNA-Seq data (Fig. S2).
Global impact of X. phaseoli pv. phaseoli on the transcriptome of common bean
The susceptible genotype initiated a more intense and diverse biological response than the resistant genotype. Indeed, differential expression analysis identified 2576 and 4503 differentially-expressed genes (DEGs) in BAT93 and JaloEEP558 respectively, which represents 12 and 22% of the total homologs, respectively (Fig. 2). Three groups of DEGs could be defined (Fig. 3). First, the core transcriptome consisting of 1482 genes simultaneously induced (758) or repressed (724) in both genotypes. Then, two specific transcriptomes, consisting of 1094 genes specifically induced (291) or repressed (803) in BAT93, and of 3021 genes specifically induced (1367) or repressed (1654) in JaloEEP558. Enrichment tests identified 83, 32 and 126 gene ontology (GO) terms enriched in the core, the BAT93-specific and the JaloEEP558-specific transcriptomes, respectively (Table S4). To remove redundancy and poorly-informative GO terms, we focused our analysis on biological process GO terms summarized using REVIGO [58]. This analysis highlighted 20, 11 and 29 GO terms enriched in the core, the BAT93-specific and the JaloEEP558-specific transcriptomes, respectively (Fig. 4). We hypothesized that the core transcriptome was representative of the general response of common bean to X. phaseoli pv. phaseoli, while the differences observed between the two specific transcriptomes likely reflected the differences in resistance or susceptibility observed between both genotypes faced to X. phaseoli pv. phaseoli.
Fig. 2.
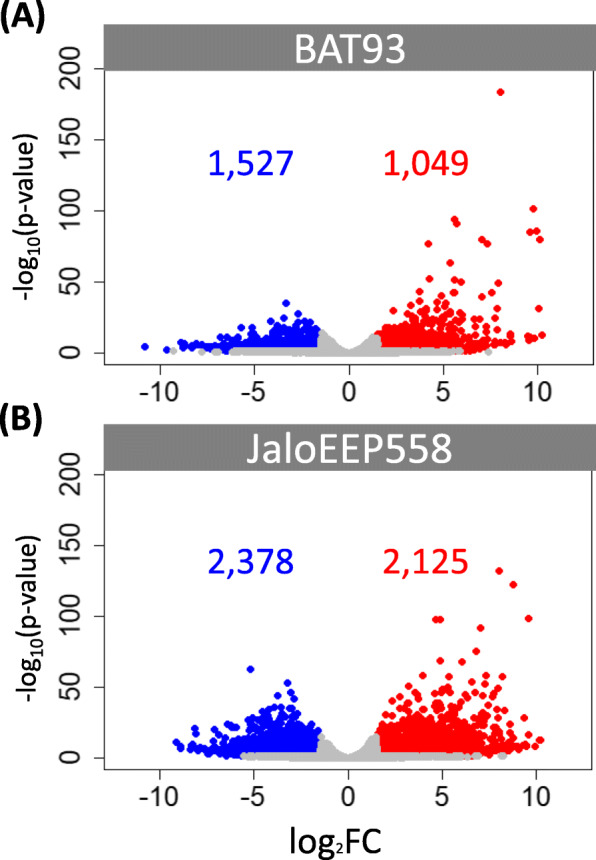
Global impact of strain CFBP6546R on the transcriptomes of BAT93 and JaloEEP558. Volcano plots represent the distribution of DEGs (adjusted p-value < 0.05, |log2FC| > 1.5) 2 days after inoculation of CFBP6546R on BAT93 (a) and JaloEEP558 (b). Each dot represents a gene either being upregulated (red), downregulated (blue) or non-differentially expressed (grey)
Fig. 3.
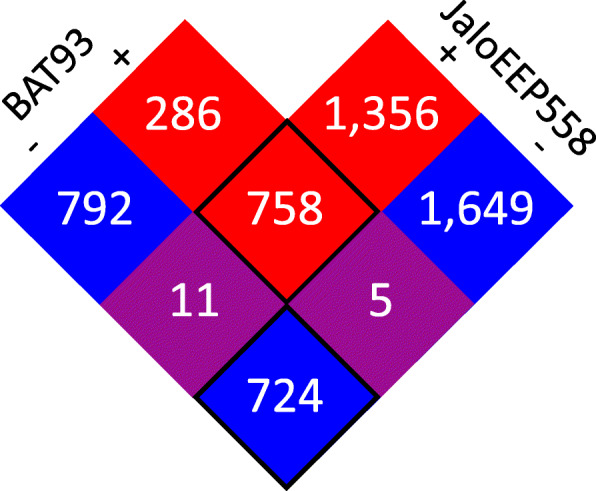
Venn diagram depicting the numbers of DEGs in BAT93 and JaloEEP558 during the interaction with strain CFBP6545R. Induced genes are highlighted in red while repressed genes are in blue. Purple squares represent genes with opposite responses in BAT93 and JaloEEP558. The core transcriptome corresponding to genes simultaneously induced or repressed in both genotypes is framed in black
Fig. 4.

Enrichment tests on the core, the BAT93-specific and the JaloEEP558-specific transcriptomes. Enrichment tests were performed using the Parametric Analysis of Gene set Enrichment (PAGE) analysis on AgriGO v2 [59]. GO terms were considered enriched when comprising a minimum of 10 genes and presenting a False Discovery Rate (FDR) below 0.05. Enriched GO terms presented here were summarized using REVIGO [58]. Mean log2FC corresponds to the mean expression calculated using the log2FC from all DEGs within each GO. Grey boxes correspond to non-enriched GO terms
Resistance was linked to downregulation of photosynthesis while susceptibility was linked to downregulation of defenses. In BAT93, the most significantly enriched GO terms were related to photosynthesis (Table S4), with a large majority of repressed genes (Fig. 4), suggesting that the resistant genotype strongly suppresses production of primary energy. On the other hand, the most significantly enriched GO terms in JaloEEP558 were related to cell death (Table S4), with a majority of repressed genes annotated as R genes from the NLR family. Together with the fact that genes belonging to the GO term “defense responses” were predominantly repressed in this genotype (Fig. 4), this suggests that global defense responses are suppressed in the susceptible genotype. Interestingly, “signaling” was enriched in the specific transcriptomes of both BAT93 and JaloEEP558 as well as in the core transcriptome. DEGs belonging to “signaling” were mostly upregulated in the core transcriptome, while specific transcriptomes comprised both upregulated and downregulated genes. This suggests that common signaling pathways were induced in both genotypes, while other were specifically up- or downregulated in one genotype or another.
GO analysis done using up- and downregulated genes separately highlighted different GO terms than when using the whole dataset of DEGs (Table S5). It pointed out an upregulation of RNA metabolism and gene expression, as well as in nitrogen compound metabolic process in JaloEEP558, while the later was downregulated in BAT93.
Detailed differences between resistant and susceptible genotypes
MapMan visualization gave a detailed overview of the differences between the resistant and susceptible genotypes (Fig. 5). In accordance with the GO analysis, Wilcoxon Rank Sum Test obtained with MapMan showed that genes related to photosynthesis were enriched in the BAT93-specific transcriptome, while defense genes related to biotic stress and signaling (RLK and PR genes) were enriched in the JaloEEP558-specific transcriptome (Table S6). Additionally, genes related to cell wall modification, ethylene signaling pathway and fatty acid metabolism were specifically enriched in JaloEEP558. To analyze the differences between both genotypes in more detail, we generated lists of genes from different classes (see Materials and Methods) and performed a KEGG analysis of hormonal signaling pathways (Table S7, Fig. S3).
Fig. 5.

MapMan overview of DEGs in metabolic pathways (a) and the photosynthetic electron transport chain (b). DEGs from the specific transcriptomes of BAT93 (left) or JaloEEP558 (right) are represented by squares colored in blue (repressed) or red (induced) following the scale bar displaying changes in gene expression values in log2FC (in the center)
NLR and RLK genes
We observed a specific repression of 30 NLR genes in the susceptible genotype while only one was specifically repressed in BAT93 (Table S7). Additionally, two NLR genes were specifically induced in JaloEEP558, while one (Phvul.006G056500) was both induced in BAT93 and repressed in JaloEEP558, thus appearing as a good candidate for being involved in CBB resistance. The RLK family presented a more complex pattern than NLRs, with a large number of RLK genes specifically induced or repressed in each genotype. More RLKs were repressed than induced in both genotypes, with 11 and 40 specifically induced and 34 and 125 specifically repressed in BAT93 and JaloEEP558, respectively (Table S7). As a result, more RLKs were differentially-expressed in the susceptible genotype than in the resistant genotype.
Kinases
A majority of kinases other than RLKs was induced in the core transcriptome (15/17, Table S7). In particular, four genes encoding calcium-dependent protein kinases (CDPKs) and calcineurin B-like protein-interacting protein kinases (CIPKs) were induced in both genotypes, indicating that genes linked to calcium signaling were induced in common bean during the interaction with X. phaseoli pv. phaseoli whatever the outcome (resistance or disease). In the JaloEEP558-specific transcriptome, a large modulation of the expression of kinases occurred, which followed the same trend of induction (20/29) than what was observed in the core transcriptome. On the other hand, BAT93-specific transcriptome was less impacted and a majority of kinases (10/12) was repressed.
Transcription factors
A larger induction of transcription factors (TFs) was specifically observed in JaloEEP558 (137/268) compared to BAT93 (28/91, Table S7). TFs from the APETALA2/ethylene response factor (AP2/ERF) superfamily presented the most differences between both genotypes (Fig. S3). Indeed, much more AP2/ERF genes were specifically induced in JaloEEP558 (32) than in BAT93 (4). In the meantime, less AP2/ERF genes were repressed in JaloEEP558 (6) than in BAT93 (8). This indicates a specific induction of AP2/ERF genes in the susceptible genotype 48 h post-inoculation. Similar patterns were observed for the MYB family, although with smaller differences between both genotypes compared to AP2/ERFs (Fig. S3). Other major TF families such as WRKY and GRAS were more impacted (i.e. both more induced and more repressed) in JaloEEP558 than in BAT93.
Lipid metabolism
Lipid metabolism was more impacted in the susceptible genotype (72 specific DEGs) than in the resistant genotype (23 specific DEGs, Table S7). Significantly, a large majority of genes involved in fatty acid synthesis (20/21) was specifically repressed in JaloEEP558 (Fig. 5a, Table S7). Some genes encoding proteins involved in lipid degradation, such as Alpha/beta hydrolases were also repressed in JaloEEP558, while others were upregulated, indicating that lipid degradation was impacted in different ways in the susceptible genotype [60]. On the other hand, fatty acid desaturases were mostly repressed in BAT93. Notably, the homolog of fatty acid desaturase 8 (FAD8, Phvul.006G068600) was both induced in JaloEEP558 and repressed in BAT93. These enzymes could play a role in defense activation [61, 62], hormone synthesis [63], increased membrane fluidity and influence membrane properties of chloroplasts [64]. Here, repression of these genes in the resistant genotype could be linked to the induction of plant defenses.
Photosynthesis and sugar metabolism
A global downregulation of photosynthesis and sugar metabolism occured in the resistant compared to the susceptible genotype (Fig. 5a). In particular, genes linked to the biosynthesis of photosystems I and II, the NAD(P) H deshydrogenase and the ATP synthase where largely repressed in BAT93 (Fig. 5b). By contrast, in JaloEEP558 three genes linked to the photosynthetic electron transport chain were induced. This result indicated a specific repression of the photosynthetic electron transport chain in the resistant genotype. More generally, the whole reactions taking place in the chloroplast appeared down-regulated in the resistant genotype, including the Calvin cycle, tetrapyrrole synthesis, photorespiration, the ascrobate and glutathione redox pathways (Fig. 5a). Sucrose and starch biosynthesis and degradation, and glygolysis were also specifically repressed in BAT93 (Fig. 5a).
Cell wall
Susceptibility was linked to the induction of cell wall modification and repression of cellulose biosynthesis while resistance was linked to the repression of genes involved in cell wall modification. Indeed, most genes involved in cell wall modification and degradation were induced in JaloEEP558 and repressed in BAT93. This was the case of xyloglucan endotransglycosylases, expansins, pectin methylesterase inhibitors, and glycosyl hydrolases (Fig. 5a and Table S7). On the other hand, genes involved in cellulose biosynthesis such as cellulose synthases were repressed in JaloEEP558.
Hormone signal transduction
Resistance and susceptibility were marked by opposite hormone signal transduction pathways. Indeed, resistance was linked to upregulation of SA signaling, while susceptibility was linked to upregulation of the ethylene pathway and downregulation of the SA and cytokinin pathways (Fig. S4). Several genes involved in resistance signaling were specifically repressed in JaloEEP558 such as the homolog of PATHOGENESIS-RELATED 1 (PR1, Phvul.006G197100) that was also specifically induced in BAT93 [65]. PR1 being a marker of SA accumulation, these results suggest that upregulation of the SA pathway occurred in the resistant genotype while it was suppressed in a susceptible context. Consistently, cytokinin signaling was suppressed in JaloEEP558. This is highlighted by the strong induction of a homolog of genes encoding type A Arabidopsis response regulators (A-ARR), which are negative regulators of cytokinin signaling and SA-dependent basal immunity [66]. As described in the MapMan enrichment analysis, many genes involved in ethylene signaling were specifically induced in the susceptible genotype, including AP2/ERF genes (Table S6, Fig. S3). This trend is confirmed by the induction of homologs from ETHYLENE RESPONSE (ETR) and ERF1/2 genes (Fig. S4). Genes involved in other hormonal signaling pathways (auxin, brassinosteroids, gibberellins or abscisic acid) also showed contrasting expression profiles between resistant and sensitive genotypes, although the differences observed were not strong enough to be conclusive.
Candidate genes for resistance to CBB
To search for genes putatively responsible for the resistance observed in BAT93, we have cross-checked different pieces of information that may suggest the involvement of certain genes in resistance to CBB. The different criteria we searched for were (i) that there was a large difference of log2FC values between the susceptible and resistant genotypes, (ii) that the gene was induced in one genotype and repressed in the other, and (iii) that the gene colocalized with a locus of resistance to CBB.
Genes with high log2FC difference
Interestingly, 201 DEGs had a large difference of expression (|Δlog2FC| > 5) between BAT93 and JaloEEP558 (Table S8). Among those genes, 148 (74%) were more expressed in JaloEEP558 than in BAT93. This proportion was higher than what was observed for the whole dataset (52%), and this difference was even higher when focusing on the top 50 genes with the largest |Δlog2FC|, among which 46 had a higher log2FC in JaloEEP558 than in BAT93 (Table S3). This indicates that the greatest differences observed between the two genotypes were due to genes strongly induced in JaloEEP558 and/or strongly repressed in BAT93. Of these 148 genes, the Heat Shock Protein (HSP) family was the most represented with 13 members, four of which belonged to the top 10 genes more expressed in JaloEEP558 than in BAT93. Significantly, three genes from the Lateral Organ Boundaries (LOB) family belonged to the list of 148 genes. Two of these LOB genes (Phvul.007G195100 and Phvul.008G257400) were ranked second and 15th of the most induced genes in JaloEEP558, suggesting that the induction of LOB genes is important for susceptibility. On the other hand, 53 genes had a higher log2FC in BAT93 than in JaloEEP558 (Table S8). A large proportion of these genes belonged to families related to resistance such as RLKs (7), NLRs (2) and PRs (2), indicating that a classical resistance response occurred in BAT93.
Genes with opposite reactions to X. phaseoli pv. phaseoli
Only 16 DEGs were simultaneously induced in one genotype and repressed in the other one (Table S9). Among those, 11 were both repressed in BAT93 and induced in JaloEEP558, suggesting that they contribute to either suppressing defenses and/or promoting disease (Fig. 3). Of those, six had also a |Δlog2FC| > 5, which encoded different proteins including a plasmodesmata-located protein (Phvul.001G229800), an HSP (Phvul.001G039700), a Lipid-Transfer Protein (Phvul.008G137100), a kinase (Phvul.008G081300), a pectin methylesterase inhibitor (Phvul.002G318500) and a sulfite exporter from the TauE/SafE family (Phvul.001G061000) (Table 2, Fig. 6). On the other hand, five genes were both induced in BAT93 and repressed in JaloEEP558, suggesting that they positively regulate resistance to CBB. Four of these genes had also a |Δlog2FC| > 5, including a placenta-specific 8 (PLAC8) family gene (Phvul.003G265800), an S-ribonuclease binding protein (SBP) family gene (Phvul.009G119200), a PR gene (Phvul.006G197100), and an ovate family gene (Phvul.009G057100).
Table 2.
Candidate genes for resistance to CBB. Grey cases indicate non differentially-expressed genes (i.e. genes with −1.5 < log2FC < 1.5 and/or adjusted p-value ≥0.05)
| Gene ID | Annotation | log2FC BAT93 | log2FC JaloEEP558 | |Δlog2FC| > 5 | Inverse pattern of expression | Colocation with QRLs |
|---|---|---|---|---|---|---|
| Phvul.001G039700 | 17.6 kDa class II HSP | -1,67 | 8,75 | Yes | Yes | – |
| Phvul.001G061000 | Sulfite exporter TauE/SafE family | -1,86 | 3,23 | Yes | Yes | – |
| Phvul.001G229800 | plasmodesmata-located protein 7 | -5,58 | 5,80 | Yes | Yes | – |
| Phvul.002G212000 | HSP20-like chaperones superfamily | -2,96 | 7,52 | Yes | – | Yes |
| Phvul.002G231500 | HSP20-like chaperones superfamily | -3,20 | 9,04 | Yes | – | Yes |
| Phvul.002G243500 | GDSL-like Lipase/Acylhydrolase superfamily | -8,35 | -2,79 | Yes | – | Yes |
| Phvul.002G249800 | RmlC-like cupins superfamily | -3,81 | 1,43 | Yes | – | Yes |
| Phvul.002G318500 | Pectin methylesterase inhibitor | -1,86 | 3,43 | Yes | Yes | – |
| Phvul.003G265800 | PLAC8 family protein | 3,25 | -4,07 | Yes | Yes | – |
| Phvul.005G014200 | NLR | -1,02 | -8,31 | Yes | – | Yes |
| Phvul.005G034000 | cytokinin oxidase/dehydrogenase 6 | −9,67 | −0,86 | Yes | – | Yes |
| Phvul.005G044600 | NAC (No Apical Meristem) domain transcriptional regulator superfamily | 0,46 | 5,50 | Yes | – | Yes |
| Phvul.005G049700 | LOB 4 | −1,25 | 4,23 | Yes | – | Yes |
| Phvul.005G058600 | MLP-like protein 43 | 4,19 | −2,73 | Yes | – | Yes |
| Phvul.005G085800 | light-harvesting chlorophyll B-binding protein 3 | −5,41 | −0,30 | Yes | – | Yes |
| Phvul.005G097600 | plasmodesmata callose-binding protein 5 | 1,33 | −3,99 | Yes | – | Yes |
| Phvul.005G097800 | bZIP transcription factor | −1,98 | 3,92 | Yes | – | Yes |
| Phvul.006G197100 | Pathogenesis-related 1 protein | 4,85 | −1,52 | Yes | Yes | – |
| Phvul.007G013000 | Eukaryotic aspartyl protease family | −5,87 | 1,45 | Yes | – | Yes |
| Phvul.007G018600 | FAD/NAD(P)-binding oxidoreductase family | −1,88 | 2,98 | – | Yes | Yes |
| Phvul.007G024800 | Major facilitator superfamily | 1,05 | −4,98 | Yes | – | Yes |
| Phvul.007G030300 | receptor serine/threonine kinase, putative | −1,32 | −6,34 | Yes | – | Yes |
| Phvul.007G038400 | CYCLIN D3;1 | −7,76 | −2,01 | Yes | – | Yes |
| Phvul.007G049700 | cysteine-rich RLK 29 | −1,88 | 1,68 | – | Yes | Yes |
| Phvul.007G051300 | cysteine-rich RLK 25 | 0,49 | −5,54 | Yes | – | Yes |
| Phvul.008G081300 | Protein kinase superfamily | −3,96 | 3,96 | Yes | Yes | – |
| Phvul.008G137100 | Lipid-transfer protein | −3,68 | 5,63 | Yes | Yes | – |
| Phvul.009G030900 | Unknown protein | −5,53 | 0,56 | Yes | – | Yes |
| Phvul.009G057100 | ovate family protein 7 | 4,64 | −4,99 | Yes | Yes | Yes |
| Phvul.009G080200 | 17.6 kDa class II HSP | −3,10 | 4,47 | Yes | – | Yes |
| Phvul.009G084400 | Integrase-type DNA-binding superfamily | −0,53 | 7,55 | Yes | – | Yes |
| Phvul.009G094000 | plant natriuretic peptide A | −0,53 | 8,66 | Yes | – | Yes |
| Phvul.009G097500 | mitotic-like cyclin 3B | −5,85 | −0,65 | Yes | – | Yes |
| Phvul.009G106200 | proline extensin-like receptor kinase 1 | −2,56 | 2,82 | Yes | – | Yes |
| Phvul.009G119200 | SBP family | 1,54 | −4,90 | Yes | Yes | – |
Fig. 6.
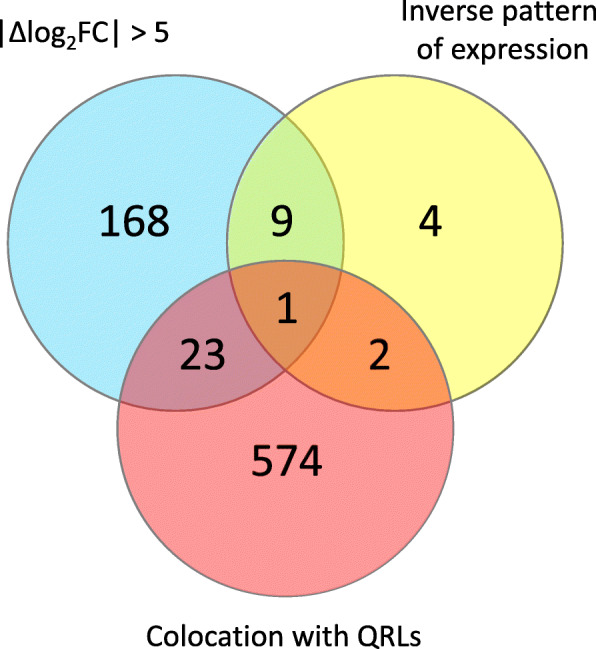
Venn diagram showing the numbers of DEGs potentially linked to resistance to CBB. The blue circle represents DEGs with a high Δlog2FC between BAT93 and JaloEEP558 (|Δlog2FC| > 5), the yellow circle represents DEGs with an inverse expression pattern between BAT93 and JaloEEP558 and the red circle represents DEGs that co-located with one of the four QRLs against CBB, described in the progeny of BAT93xJaloEEP558 [50]
Genes within QRLs
We positioned the four CBB QRLs previously identified in BAT93 on the bean reference genome and extracted the DEGs from specific transcriptomes located within these putative QRLs (Table S10). A total of 600 DEGs were retrieved in these four CBB QRLs, among which 24 (4.0%) had a |Δlog2FC| > 5, which is similar to the ratio observed in the whole transcriptomic data where 201 out of 5581 DEGs (3.6%) had a |Δlog2FC| > 5 (Fig. 6). In addition, 12 and 21% of the total genes were differently expressed in the QRLs of BAT93 and JaloEEP558, respectively, which is not different from what was observed for the whole transcriptome (Fig. 2). Thus, no specific pattern of expression could be observed in the QRLs compared to the rest of the common bean genome. Among the genes in QRLs with |Δlog2FC| > 5, the three most repressed genes in JaloEEP558 were one NLR gene (Phvul.005G014200) and two RLK genes (Phvul.007G051300 and Phvul.007G030300). Thus, these three genes represent good candidates for CBB resistance. The previously described HSP (Phvul.002G231500, Phvul.002G212000 and Phvul.009G080200) and LOB (Phvul.005G049700) genes specifically induced in JaloEEP558 or repressed in BAT93 were also retrieved in these QRLs.
Only one gene presented the three characteristics of being induced in BAT93 while repressed in JaloEEP558, having a |Δlog2FC| > 5 and locating in a QRL to CBB. This gene (Phvul.009G057100) is located within the QRL associated to marker D0157 on chromosome 9. It belongs to the ovate family, which is involved in plant growth regulation and can suppress elongation [67–69].
Discussion
In this work, we used RNA-Seq data to investigate the transcriptomic response of resistant and susceptible common bean genotypes during their interaction with X. phaseoli pv. phaseoli. Importantly, our work provided novel whole genome sequence data of two parental lines of a reference population (BAT93 x JaloEEP558) used in many genetic and genomic studies, including mapping of disease resistances [70–78].
A global trend was that the proportion of DEGs was higher in the susceptible genotype (22%) than in the resistant genotype (12%). This result highlights that susceptibility results in a larger reprogramming of gene expression than resistance, which was not surprising since a similar trend was observed in many other studies describing plant transcriptomic responses to diverse plant pathogens including fungi, bacteria and viruses [79–83].
In the core transcriptome, the induction of genes from the CIPK and CDPK families suggests that both genotypes are able to perceive X. phaseoli pv. phaseoli through Ca2+ signaling [84–87]. Following this hypothesis, the susceptibility to CBB observed in JaloEEP558 would result from inhibition of PTI by bacterial effectors rather than from non-detection by PRRs. In accordance with this, the susceptible genotype displayed a large repression of genes putatively involved in PTI such as RLKs, as well as defense response and NLR genes. This indicates that successful bean colonization by X. phaseoli pv. phaseoli is linked to suppression of plant defenses and reflects a potential involvement of bacterial effectors.
Pathogenic bacteria from the genus Xanthomonas are often described as hemibiotrophic [88]. Here, apparition of necrotic tissues suggested that X. phaseoli pv. phaseoli shifted from biotrophic to necrotrophic from 8 to 15 DPI. The defense response to biotrophic and hemibiotrophic pathogens is usually regulated by SA while defense responses to necrotrophic pathogens classically involve JA and ethylene [13, 89]. Here, a global induction of the SA pathway was linked to resistance, suggesting that an adapted SA response is effective in BAT93 48 h after infection by X. phaseoli pv. phaseoli. On the other hand, susceptibility was linked to the induction of the ethylene pathway and repression of the SA pathway, which are often described as being antagonistic to each other [89]. The induction of the ethylene pathway is reminiscent of the observation that ethylene is implicated in increased symptoms in other Xanthomonas-plant pathosystems [90–92]. In this view, it is tempting to speculate that ethylene is involved in successful colonization by X. phaseoli pv. phaseoli, while SA is involved in common bean resistance to CBB.
The specific suppression of photosynthesis, sugar metabolism and other chloroplast-associated genes observed in BAT93 reflects a rather classical defense response. Indeed, similar trends have been described using transcriptomics in different interactions between plants and bacteria [93–96] and integration of transcriptomic data from different pathosystems led to the hypothesis that suppression of photosynthesis is part of the plant adaptive immunity [97]. In addition, several studies have shown that incompatible interaction is linked to a decrease in photosynthetic activity [98–101] and that inhibition of photosynthesis often leads to the accumulation of reactive oxygen species [102]. Supporting this, it is interesting to note that Xanthomonas citri pv. citri is able to counter the decrease in photosynthesis by mimicking a plant natriuretic peptide, leading to suppression of resistance in citrus leaves [103].
The plant cell wall plays an important role in plant immunity, both as a physical barrier against pathogens and by releasing signaling compounds known as DAMPs when altered [104]. Successful pathogens are usually able to degrade the plant cell wall to access nutrients. In JaloEEP558, specific induction of genes involved in cell wall modification, such as expansins, xyloglucan endotransglycosylases and glycosyl hydrolases, suggests that remodeling of cell wall occurred in the susceptible genotype [105, 106]. In accordance with this, early CBB phenotype usually corresponds to water-soaking symptoms involving the softening and loosening of the cell wall. In contrast, cell wall modification genes were repressed in BAT93, which could suggest that cell wall rigidification occurred in the resistant genotype, thus preventing bacterial progression. Modification of the cell wall is tightly linked to cell size and shape and to morphogenesis of plant organs [107]. Interestingly, transcription factors from the LOB and ovate families were found both among the most differentially expressed genes between BAT93 and JaloEEP558, and among the DEGs located in QRLs to CBB.
LOB is a family of plant-specific transcription factors with key roles in the regulation of plant organ development [108, 109]. Induction of CsLOB1 by Xanthomonas in Citrus sinensis triggers cell expansion and is required for symptom development [57]. Induction of LOB genes can induce genes involved in cell wall modification such as pectin methylesterase inhibitors [110]. Here, the strong induction of LOB genes in JaloEEP558 could contribute to the induction of downstream genes involved in cell wall modification and to the development of symptoms. The LOB gene Phvul.005G049700 is located on chromosome 5 and colocates with a QRL linked to marker D1081, explaining 15% of the phenotypic vartiation [50]. Therefore, Phvul.005G049700 could putatively act as a negative regulator of resistance to CBB.
On the other hand, the ovate family member Phvul.009G057100 was strongly induced in the resistant genotype and repressed in the susceptible genotype. This gene could positively contribute to CBB resistance as it colocates with a QRL on chromosome 9 that is linked to marker D0157, and explains 13% of the phenotypic variation [50]. Consistent with this hypothesis, ovate family members act as transcriptional repressors involved in the suppression of cell growth and elongation as well as in the regulation of secondary cell wall and vascular development [67–69]. However, to our knowledge, the ovate family has not so far been described as playing any role in plant-pathogen interaction.
Altogether, our analyses pointed out different molecular pathways appearing important for either promoting disease in the susceptible genotype or triggering immunity in the resistant genotype. The genes involved in these pathways were scattered throughout the whole common bean genome, which reflects the complexity of CBB resistance. In particular, large clusters of dozens of NLR genes exist at common bean subtelomeres [111–114] but the 30 NLRs repressed in JaloEEP558 following X. phaseoli pv. phaseoli infection did not correspond to the specific repression of one of these clusters. Thus, no particular locus was unveiled as being responsible for CBB resistance. To summarize, resistance was linked to suppression of photosynthesis and sugar metabolism and induction of the SA pathway, while susceptibility was linked to downregulation of plant defenses and upregulation of the ethylene pathway and AP2/ERF transcription factors as well as genes involved in cell wall modification.
Xanthomonas bacteria possess transcription activator-like effectors (TALE) that are type III effectors able to induce the transcription of genes by specifically binding to the promoter of plant susceptibility genes and recruiting the transcription machinery [115]. Nine different TALE-encoding genes and alleles have been described in CBB agents [116]. Strain CFBP6546R used in this study bears tal19I and tal18H [116]. Interestingly, most of the pathways induced in the susceptible genotype or repressed in the resistant genotype were previously described as being induced by TALEs to promote disease [117]. For example, the best-characterized TALE targets so far are SWEET genes that encode sugar exporters presumably providing nutrients for the pathogen [118–121]. SWEET gene induction by TALE has been described in the interaction of Xanthomonas with rice [121–127] cassava [128] and cotton [129]. According to our results, it is tempting to speculate that the suppression of photosynthesis and sugar metabolism observed in the resistant genotype could lead to the reduction of sugar production by the plant cells, thus contributing to resistance by depriving bacteria of sugar. Interestingly, one SWEET gene (Phvul.009G134300) was specifically repressed in the resistant genotype, while another one (Phvul.002G203600) was specifically induced in the susceptible genotype. However, Phvul.002G203600 was not predicted as a target of TAL19I or TAL18H [116]. Thus, but it seems that its induction was not due to the action of TALEs.
Other TALE targets include different types of TFs from the AP2/ERF [130], bHLH [54, 131] or LOB [57, 132, 133] families, which is reminiscent of what was observed here in the susceptible genotype. Interestingly, these targets are often linked to cell wall reorganization and modification of the plant cells shape. In pepper, the TALE AvrBS3 from Xanthomonas campestris pv. vesicatoria induces the bHLH TF UPA20, which leads to the hypertrophy of leaf cells [131]. In tomato, AvrHah1 from X. gardneri targets another bHLH TF whose induction upregulates the expression of a pectate lyase responsible for the apparition of water soaking symptoms [54]. In citrus, the induction of CsLOB1 by different TALEs from X. citri pv. citri or X. citri pv. aurantifolii is required for the apparition of canker symptoms due to hyperplasia and rupture of the epidermis in infected tissue [57, 132, 133]. Here, the JaloEEP558-specific induction of LOB homologs and other genes involved in cell wall degradation and modification suggests that X. phaseoli pv. phaseoli employs mechanisms similar to what was observed in tomato and citrus. The parallel observed between our transcriptomic data and TALE targets in different plant species suggests that TALE evolution was driven by the necessity to target pre-existing susceptibility hubs in plants. This also raises the question as to whether AP2/ERF, bHLH, pectate lyases, or LOB homologs could constitute direct targets for TALEs from X. phaseoli pv. phaseoli in common bean. Thus, searching for X. phaseoli pv. phaseoli TALE targets in common bean would bring valuable knowledge on the molecular interactions between common bean and X. phaseoli pv. phaseoli.
Conclusions
The analyses presented here bring novel information about the transcriptomic response of common bean to X. phaseoli pv. phaseoli attack, and may help to better understand the mechanisms underlying resistance to CBB in common bean. The global trends observed here lead to the hypothesis that the transcriptome of susceptible varieties is largely manipulated by the bacterium, while resistant varieties retain control of their transcriptome, increase signaling and adapt their metabolism for defense purposes.
Methods
Plant materials, bacterial strains and growing conditions
The original seeds from common bean cultivars BAT93 (Mesoamerican) and JaloEEP558 (Andean) were obtained from the Center for Tropical Agriculture (CIAT, Colombia) and are available under accession numbers G51294 and G9603, respectively (http://genebank.ciat.cgiar.org/genebank/main.do). Common bean cultivars were seeded in plastic pots (7 × 7 × 8 cm) containing pre-wetted soil. Plants were grown in a growth chamber at 23 °C/20 °C (day/night) with 80% relative humidity and a photoperiod of 16 h. Plants were watered every 2 days with water for the first 10 days, then with a nutrient solution of N-P-K (15–10-30). The day before inoculation, relative humidity and temperature were increased at 95% and 28 °C/25 °C (day/night) to provide adequate conditions for infection.
X. phaseoli pv. phaseoli strain CFBP6546R is a rifamycin-resistant derivative of strain CFBP6546. This strain was grown at 28 °C for 48 h on Trypticase Soy Agar (TSA) medium (17.0 g.L− 1 pancreatic digest of casein; 3.0 g.L− 1 enzymatic digest of soya bean; 5.0 g.L− 1 NaCl; 2.5 g.L− 1 K2HPO4; 2.5 g.L− 1 glucose; 15 g.L− 1 agar) supplemented by rifamycin (50 mg. L− 1), then at 28 °C for 24 h on TSA10 (a 1/10 dilution of TSA, except for agar maintained at 15 g.L− 1) supplemented by rifamycin (50 mg. L− 1) to obtain fresh bacterial cultures.
Pathogenicity assays
Inoculations were performed at stage V1 (first trifoliolate leaf unfolded) by dipping the first trifoliate leaf for 30 s into bacterial suspensions calibrated at 1 × 107 CFU.mL− 1 in sterile distilled water (CFU: colony-forming unit). Water-inoculated plants were used as control. For bacterial population sizes, at least three plants per condition were harvested at 2 h, 1, 5, 8, and 15 days post-inoculation (DPI). Each trifoliate leaf was ground individually for 2 min in a plastic bag supplemented with 10 mL of distilled water, using a Stomacher 80 (Seward, London, United Kingdom) at maximum power. Appropriate dilutions were plated on TSA10 supplemented by rifamycin (50 mg. L− 1) and incubated at 28 °C for 72 h before counting. At 8 and 15 DPI, symptom development was monitored by chlorophyll fluorescence imaging [134] at the PHENOTIC Seeds and Plants platform of the IRHS in Angers (France). Each leaflet was set in the dark for 30 min. Then, a first picture was taken under a modulated flash of light to measure basal fluorescence (F0) of the tissues, followed by another picture taken under a high flash of saturating light to measure the maximum fluorescence emission level (Fm). For each pixel, the maximum quantum yield of photosystem II photochemistry (Fv/Fm = (Fm-F0)/Fm) was calculated with Phenoplant (http://www.phenoplant.org) to discriminate diseased and healthy leaf areas [52]. Diseased areas were divided into different Fv/Fm clusters according to the intensity of the symptoms (impacted, wilted and necrotic) [52]. Pathogenicity assays were performed twice independently.
Bean whole genome sequencing, assembly and annotation
Genomic DNA was extracted from young leaves and buds using the Nucleospin Plant II kit (Macherey-Nagel, Hoerdt, France). Paired libraries were prepared using the Illumina TruSeq Nano DNA Library prep kit, then sequenced at 2 × 150 bp on an Illumina HiSeq3000. Reads were trimmed to the first undefined base and assembled using SOAPdenovo software with k-mer size of 81 bp and max read length of 100 bp. Assembled contigs were gap filled and spurious assemblies were removed using redundancy search (97% identity, maximum overhang 50 bp). Scaffolds of minimum 1000 bp with at least 500 bp defined were retained. Annotation of the whole genome assembly was performed using EuGene-EP pipeline [135] using our BAT93 or JaloEEP558 RNA-Seq assemblies and four different protein databases as training sets for structural annotation: TAIR10, Swiss-Prot, UniProt plant subset and predicted proteins from the soybean (Glycine max) reference genome [136]. Gene completeness was assessed using BUSCO version 1.22 and the plantae dataset [137]. Gene functions presented here were determined by BLASTp (e-value ≤1 × 10− 6) on the P. vulgaris v2.1 reference genome from genotype G19833 [18]. Homologs between BAT93 and JaloEEP558 were searched for by reciprocal BLASTp (e-value ≤1 × 10− 6) of all predicted genes, and keeping only genes having the same best hit after both reciprocal BLASTp searches and the same best hit after BLASTp on the P. vulgaris reference genome. The four QRLs to CBB described in a BAT93 x JaloEEP558 progeny [50] were delimited by using the sequence of restriction fragment length polymorphism (RFLP) markers flanking these QRLs (Table S11), available at the PhaseolusGenes database (http://phaseolusgenes.bioinformatics.ucdavis.edu).
RNA-Seq experiments
In order to obtain tissues with homogeneous and synchronous plant cells interacting with bacteria, leaflets from the first trifoliate leaves at stage V1 were detached from the plant, and vacuum-infiltrated for 2 × 1 min into a bacterial suspension of CFBP6546R strain at 1 × 108 CFU.mL− 1 diluted in sterile distilled water, or pure sterile distilled water as control. Infiltrated leaflets were maintained in Petri dishes by dipping the petiole in water agar (0.7%) and incubated at 28 °C/25 °C (day/night) with a photoperiod of 16 h. Fourty-eigth hours after inoculation, 10 disks were sampled with a 1.3 cm diameter borer and immediately frozen in liquid nitrogen. Frozen disks were ground using a ball mill for 30 s at 25 Hz. Total RNA was extracted with the TRIzol® Plus RNA Purification Kit (Ambion, Applied Biosystems, Courtaboeuf, France) according to manufacturer’s recommendations, except that TRIzol® was used at 60 °C and that an additional DNase treatment was performed. The quantity and quality of total DNA-free RNA were evaluated using a NanoDropTM (Thermo Scientific, Wilmington, USA) and an ExperionTM chip (Bio-Rad, Hercules, CA, USA). Then, for each modality, RNA-Seq was performed on three independent biological replicates at the GeT-PlaGe core facility, INRAE Toulouse. RNA-Seq libraries have been prepared according to Illumina’s protocols using the Illumina TruSeq Stranded mRNA sample prep kit to analyze mRNA. Briefly, mRNAs were selected using poly-T beads. Then, RNAs were fragmented to generate double stranded cDNAs and adaptators were ligated to be sequenced. Eleven cycles of PCR were applied to amplify libraries. Library quality was assessed using a Fragment Analyser and libraries were quantified by qPCR using the Kapa Library Quantification Kit. All libraries were pooled and the whole pool was loaded into two sequencing lanes. RNA-Seq experiments have been performed on an Illumina HiSeq3000 using a paired-end read length of 2 × 150 pb with the Illumina HiSeq3000 sequencing kits. Total reads were mapped on the annotated genome sequences of BAT93 or JaloEEP558 and counted using the glint software (http://lipm-bioinfo.toulouse.inra.fr/download/glint). Only best-scores were taken into account, with a minimal hit length of 40 bp, a maximum of 5 mismatches and no gap allowed. Ambiguous matches with the same best score were removed. Principal component analysis and a Pearson correlation matrix were performed with R, using the number of reads per genes within each sample.
Analysis of differentially-expressed genes
Differentially-expressed genes (DEGs) were determined using DeSeq2 with default normalization settings [138] and with adjusted p-value < 0.05 [139]. Three different log2FC thresholds were tested, at |log2FC| > 1, 1.5, or 2.0. DEGs were grouped according to three expression patterns. First, the core transcriptome consisting of genes simultaneously induced or repressed in both genotypes. Then, the BAT93-specific and JaloEEP558-specific transcriptomes consisting of genes specifically induced or repressed in BAT93 or JaloEEP558, respectively. Each gene set was used to perform an enrichment test based on Gene Ontologies (GOs) using the Parametric Analysis of Gene set Enrichment (PAGE) available on AgriGO v2.0 (http://systemsbiology.cau.edu.cn/agriGOv2) [59]. GO terms represented by a minimum of 10 genes and a False Discovery Rate (FDR) < 0.05 were considered significantly enriched. Enriched GO terms belonging to biological processes were summarized using REVIGO, a computational approach that summarizes long GO lists by reducing functional redundancies (http://revigo.irb.hr) [58]. Enriched GO terms from the three different log2FC thresholds were compared to each other, either by using all DEGs (Table S12). Consistency of enriched GO terms among the different log2FC tresholds was observed. Therefore, we used the intermediate log2FC threshold of 1.5 for all analyses. Functional annotations of DEGs were visualized using MapMan (http://mapman.gabipd.org) [140]. DEGs linked to hormone signaling were visualized on the pvu04075 pathway available on KEGG mapper [141] (https://www.genome.jp/kegg/mapper.html). Lists of DEGs associated with the cell wall or the metabolism of lipids were extracted using corresponding MapMan Bin codes. Transcription factors (TFs), kinase genes and RLKs used in this study were retrieved from the iTAK database (http://itak.feilab.net) [142]. The complete set of common bean NLRs was retrieved according to Richard et al. 2017a [143].
RT-qPCR assays
To validate RNA-Seq results, RT-qPCR assays were performed on 10 selected genes representing different patterns of induction or repression in both bean genotypes. For each sample, cDNAs were synthetized from the total RNA using oligo (dT)15 primer with the M-MLV reverse transcriptase (Promega, Madison, WI, USA) according to manufacturer’s recommendations. RT-qPCR primers were designed using Primer3 [144] and checked for the absence of self or cross dimerization using Netprimer (https://www.premierbiosoft.com/netprimer, Table S13). Specificity and efficiency of each primer pair were checked by RT-qPCR and melting curves on a serial dilution of pooled cDNAs from the sample set. All primers produced a single peak with an efficiency ranging between 90 and 100%. RT-qPCR were performed with the MESA BLUE qPCR 2X MasterMix Plus for SYBR kit (Eurogentec, Seraing, Belgium) in a thermocycler with the following cycle: denaturation at 95 °C for 5 min, then 40 cycles of denaturation at 95 °C for 15 s followed by annealing and extending at 60 °C for 40 s. Relative expression levels were calculated using the 2–ΔΔCt method for each gene and were normalized to the threshold cycle (Ct) values of the internal common bean reference genes EF1-alpha (Phvul.004G060000), Actin 11 (Phvul.008G011000) and Insulin-Degrading Enzyme (IDE, Phvul.001G133200) [145]. For each modality, three technical replicates were made for each of the three biological replicates.
Supplementary information
Additional file 1: Figure S1 Statistical analysis of RNA-Seq data. Statistics were performed by using the total number of RNA-Seq reads mapped on the 20,787 homologs shared by BAT93 and JaloEEP558. Principal component analysis (A). Triangles and dots represent the barycenters of RNA-Seq data from BAT93 and JaloEEP558, respectively. Data from plants inoculated with H2O are in orange while data from plants inoculated with Xanthomonas strain CFBP6546R are in black. Pearson correlation matrix (B). Numbers represent the means of Pearson correlation coefficients calculated using three biological replicates per condition.
Additional file 2: Figure S2. Validation of RNA-Seq results by RT-qPCR analysis in BAT93 (A) and JaloEEP558 (B). Gene expression data were expressed according to the 2–ΔΔCt method (Vandesompele et al. 2002) [145], relatively to three housekeeping genes: Act11, EF1-α and IDE, and to the value of water inoculated plants.
Additional file 3 Figure S3 Numbers of DEGs in the different families of transcription factors. The list of common bean transcription factors was retrieved from the iTAK database (http://itak.feilab.net, Zheng et al. 2016) [142]. The numbers of induced and repressed genes are represented by red and blue bars respectively, in the specific transcriptomes of BAT93 (solid) or JaloEEP558 (hatched). Only families with at least 10 DEGs were represented.
Additional file 4: Figure S4. KEGG orthology map for plant hormone signal transduction. The KEGG orthology maps (pvu04075) of BAT93 (A) and JaloEEP558 (B) highlight DEGs that were induced (red), repressed (blue), or both induced and repressed (green) 48 h after inoculation with Xanthomonas phaseoli pv. phaseoli. The number of DEGs (colored numbers) and the average Log2FC (black numbers) are indicated above the boxes.
Additional file 5: Table S1. Number of genes shared among common bean genomes, as determined by reciprocal blastp (e-value <1E-6).
Additional file 6: Table S2. List of homologs shared by BAT93 and JaloEEP558.
Additional file 7: Table S3. List of DEGs in BAT93 and JaloEEP558 48 h after inoculation by X. phaseoli pv. phaseoli strain CFBP6546R.
Additional file 8: Table S4. Raw enrichment tests on the core, BAT93-specific or JaloEEP558-specific transcriptomes.
Additional file 9: Table S5. Enrichment tests on the core, BAT93-specific or JaloEEP558-specific transcriptomes, using induced or repressed DEGs separately.
Additional file 10: Table S6. List of enriched MapMan bins in the BAT93-specific and JaloEEP558-specific transcriptomes.
Additional file 12: Table S8. List of DEGs with |Δlog2FC| > 5.
Additional file 13: Table S9. List of DEGs with an inverse expression pattern between BAT93 and JaloEEP558.
Additional file 14: Table S10. List of DEGs in the four QRLs to CBB described in the progeny of BAT93xJaloEEP558 (Nodari et al., 1993) [50].
Additional file 15: Table S11. Genetic markers associated to the four QRLs to CBB described by Nodari et al. 1993 [the four QRLs to CBB described in the progeny of BAT93xJaloEEP558 (Nodari et al., 1993) [50].
Additional file 16: Table S12. Results of the enrichment tests according to the core, BAT93-specific or JaloEEP558-specific transcriptome and according to different log2FC threshold.
Additional file 17: Table S13. Primers used for RT-qPCR assays.
Acknowledgements
The authors thank Céline Rousseau from the PHENOTIC platform for her help in the analysis of chlorophyll fluorescence and Muriel Bahut from the ANAN platform for her help in performing RT-qPCR assays. The authors also thank the French Network on Xanthomonads (FNX) (https://www.reseau-xantho.org/) for recurrent scientific exchange within the framework of the ANR CROpTAL project and particularly Lionel Gagnevin for critical reading of the manuscript. Bacterial strain preservation and supply is ensured by the CIRM-CFBP (Beaucouzé, INRA, France; https://www6.inrae.fr/cirm_eng/CFBP-Plant-Associated-Bacteria).
Abbreviations
- CBB
Common bacterial blight of bean
- RNA-Seq
RNA sequencing
- TALE
Transcription activator-like effector
- PAMP/MAMP
Pathogen−/microbe-associated molecular pattern
- PRR
Pattern-recognition receptor
- RLK
Receptor-like kinase
- PTI
PAMP-triggered immunity
- ETI
Effector-triggered immunity
- R gene/protein
Resistance gene/protein
- NLR
Nucleotide-binding leucine-rich repeat
- MAPK
Mitogen-activated protein kinase
- SA
Salicylic acid
- JA
Jasmonic acid
- PR
Pathogenesis-related
- PCR
Polymerase chain reaction
- EU
European Union
- QRL
Quantitative resistance locus
- cDNA-AFLP
CDNA-amplified fragment length polymorphism
- qRT-PCR
Quantitative real-time PCR
- DPI
Days post inoculation
- DEG
Differentially-expressed gene
- GO
Gene ontology
- CDPK
Calcium-dependent protein kinase
- CIPK
Calcineurin B-like protein-interacting protein kinase
- TF
Transcription factor
- AP2/ERF
APETALA2/ethylene response factor
- bHLH
Basic Helix-Loop-Helix
- HSP
Heat-shock protein
- MLO
Mildew resistance locus
- EDS1
ENHANCED DISEASE SUSCEPTIBILITY 1
- PR1
PATHOGENESIS-RELATED 1
- JAZ1
JASMONATE ZIM-domain 1
- FAD8
Fatty acid desaturase 8
- LOB
Lateral Organ Boundaries
- PLAC8
Placenta-specific 8
- SBP
S-ribonuclease binding protein
- DAMP
Damage-associated molecular pattern
- FDR
False Discovery Rate
- ETR
ETHYLENE RESPONSE
- CIAT
Center for Tropical Agriculture
- TSA
Trypticase Soy Agar
- RFLP
Restriction fragment length polymorphism
- PAGE
Parametric Analysis of Gene set Enrichment
- CFU
Colony-forming units
- gFM
Grams of fresh materials
Authors’ contributions
JF, MR, MAJ and NWGC conceived and designed the experiments. JF, MR, RS, AP and NWGC performed the experiments. JF, MR, SC, SP, MB and NWGC analyzed the data. JF, MAJ and NWGC wrote the manuscript. All authors have read and approved the manuscript.
Funding
This study was supported by the French National Research Agency CROpTAL project to NWGC (ANR-14-CE19–0002-04). JF is funded by a PhD grant (BRUTAL project) from the French National Research Institute for Agriculture (INRAE) and the region Pays de la Loire, France. MR was funded by a PhD grant (X-HOT project) from Angers-Loire Metropole, France. This work was performed in collaboration with the GeT core facility, Toulouse, France (http://get.genotoul.fr), and was supported by France Génomique National infrastructure, funded as part of “Investissement d’avenir” program managed by Agence Nationale pour la Recherche (contract ANR-10-INBS-09). The funding bodies did not participate in any of the study design, data collection and analysis, or writing the manuscript.
Availability of data and materials
Whole genome assemblies generated for this study have been submitted to GenBank under accession numbers JAAIFG000000000 (BAT93) and JAAIFH000000000 (JaloEEP58). Sequences and annotations are available at https://bbric-pipelines.toulouse.inra.fr/myGenomeBrowser?public=1. The raw RNA-Seq data and metadata have been submitted to the Gene Expression Omnibus under the accession number GSE155080 (SRA: SRP273448).
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Justine Foucher, Email: justine.foucher@inrae.fr.
Mylène Ruh, Email: mylene.ruh@hotmail.fr.
Anne Préveaux, Email: anne.preveaux@inrae.fr.
Sébastien Carrère, Email: sebastien.carrere@inrae.fr.
Sandra Pelletier, Email: sandra.pelletier@inrae.fr.
Martial Briand, Email: martial.briand@inrae.fr.
Rémy-Félix Serre, Email: remy-felix.serre@inrae.fr.
Marie-Agnès Jacques, Email: marie-agnes.jacques@inrae.fr.
Nicolas W. G. Chen, Email: nicolas.chen@agrocampus-ouest.fr
Supplementary information
Supplementary information accompanies this paper at 10.1186/s12864-020-06972-6.
References
- 1.Jones JDG, Dangl L. The plant immune system. Nature. 2006;444:323–329. doi: 10.1038/nature05286. [DOI] [PubMed] [Google Scholar]
- 2.Dodds PN, Rathjen JP. Plant immunity: towards an integrated view of plant–pathogen interactions. Nat Rev Genet. 2010;11:539–548. doi: 10.1038/nrg2812. [DOI] [PubMed] [Google Scholar]
- 3.Boller T, Felix G. A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu Rev Plant Biol. 2009;60:379–406. doi: 10.1146/annurev.arplant.57.032905.105346. [DOI] [PubMed] [Google Scholar]
- 4.Zipfel C. Plant pattern-recognition receptors. Trends Immunol. 2014;35:345–351. doi: 10.1016/j.it.2014.05.004. [DOI] [PubMed] [Google Scholar]
- 5.Böhm H, Albert I, Fan L, Reinhard A, Nürnberger T. Immune receptor complexes at the plant cell surface. Curr Opin Plant Biol. 2014;20:47–54. doi: 10.1016/j.pbi.2014.04.007. [DOI] [PubMed] [Google Scholar]
- 6.Macho AP, Zipfel C. Plant PRRs and the activation of innate immune signaling. Mol Cell. 2014;54:263–272. doi: 10.1016/j.molcel.2014.03.028. [DOI] [PubMed] [Google Scholar]
- 7.Feng F, Zhou JM. Plant-bacterial pathogen interactions mediated by type III effectors. Curr Opin Plant Biol. 2012;15:469–476. doi: 10.1016/j.pbi.2012.03.004. [DOI] [PubMed] [Google Scholar]
- 8.Büttner D. Behind the lines-actions of bacterial type III effector proteins in plant cells. FEMS Microbiol Rev. 2016;40:894–937. doi: 10.1093/femsre/fuw026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Izoré T, Job V, Dessen A. Biogenesis, regulation, and targeting of the type III secretion system. Structure. 2011;19:603–612. doi: 10.1016/j.str.2011.03.015. [DOI] [PubMed] [Google Scholar]
- 10.Galán JE, Collmer A. Type III secretion machines: bacterial devices for protein delivery into host cells. Science. 1999;284:1322–1328. doi: 10.1126/science.284.5418.1322. [DOI] [PubMed] [Google Scholar]
- 11.Cui H, Tsuda K, Parker JE. Effector-triggered immunity: from pathogen perception to robust defense. Annu Rev Plant Biol. 2015;66:487–511. doi: 10.1146/annurev-arplant-050213-040012. [DOI] [PubMed] [Google Scholar]
- 12.Bigeard J, Colcombet J, Hirt H. Signaling mechanisms in pattern-triggered immunity (PTI) Mol Plant. 2015;8:521–539. doi: 10.1016/j.molp.2014.12.022. [DOI] [PubMed] [Google Scholar]
- 13.Bari R, Jones JDG. Role of plant hormones in plant defence responses. Plant Mol Biol. 2009;69:473–488. doi: 10.1007/s11103-008-9435-0. [DOI] [PubMed] [Google Scholar]
- 14.Pieterse CMJ, Van der Does D, Zamioudis C, Leon-Reyes A, Van Wees SCM. Hormonal modulation of plant immunity. Annu Rev Cell Dev Biol. 2012;28:489–521. doi: 10.1146/annurev-cellbio-092910-154055. [DOI] [PubMed] [Google Scholar]
- 15.Malinovsky FG, Fangel JU, Willats WGT. The role of the cell wall in plant immunity. Front Plant Sci. 2014;5:178. doi: 10.3389/fpls.2014.00178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Broughton WJ, Hernandez G, Blair M, Beebe S, Gepts P, Vanderleyden J. Beans (Phaseolus spp.) - model food legumes. Plant Soil. 2003;252:55–128. doi: 10.1023/A:1024146710611. [DOI] [Google Scholar]
- 17.Foyer CH, Lam HM, Nguyen HT, Siddique KHM, Varshney RK, Colmer TD, et al. Neglecting legumes has compromised human health and sustainable food production. Nat Plants. 2016;2:16112. doi: 10.1038/nplants.2016.112. [DOI] [PubMed] [Google Scholar]
- 18.Schmutz J, McClean PE, Mamidi S, Wu GA, Cannon SB, Grimwood J, et al. A reference genome for common bean and genome-wide analysis of dual domestications. Nat Genet. 2014;46:707–713. doi: 10.1038/ng.3008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Delgado-Salinas A, Bonet A, Gepts P. The wild relative of Phaseolus vulgaris in middle america. 1988. [Google Scholar]
- 20.Rodríguez De Luque JJ, Creamer B. Major constraints and trends for common bean production and commercialization; establishing priorities for future research. Agron Colomb. 2014;32:423–431. doi: 10.15446/agron.colomb.v32n3.46052. [DOI] [Google Scholar]
- 21.Singh SP, Miklas PN. Breeding common bean for resistance to common blight: a review. Crop Sci. 2015;55:971–984. doi: 10.2135/cropsci2014.07.0502. [DOI] [Google Scholar]
- 22.EFSA Panel on Plant Health (PLH) Scientific Opinion on the pest categorisation of Xanthomonas axonopodis pv. phaseoli and Xanthomonas fuscans subsp. fuscans. EFSA J. 2014;12:3856. doi: 10.2903/j.efsa.2014.3856. [DOI] [Google Scholar]
- 23.Belete T, Bastas K. Common bacterial blight (Xanthomonas axonopodis pv. phaseoli) of beans with special focus on Ethiopian condition. J Plant Pathol Microbiol. 2017;8:403. doi: 10.4172/2157-7471.1000403. [DOI] [Google Scholar]
- 24.Vauterin L, Hoste B, Kersters K, Swings J. Reclassification of Xanthomonas. Int J Syst Bacteriol. 1995;45:472–489. doi: 10.1099/00207713-45-3-472. [DOI] [Google Scholar]
- 25.Constantin EC, Cleenwerck I, Maes M, Baeyen S, Van Malderghem C, De Vos P, et al. Genetic characterization of strains named as Xanthomonas axonopodis pv. dieffenbachiae leads to a taxonomic revision of the X. axonopodis species complex. Plant Pathol. 2016;65:792–806. doi: 10.1111/ppa.12461. [DOI] [Google Scholar]
- 26.Oren A, Garrity GM. Notification of changes in taxonomic opinion previously published outside the IJSEM. Int J Syst Evol Microbiol. 2017;67:2081–2086. doi: 10.1099/ijsem.0.002071. [DOI] [PubMed] [Google Scholar]
- 27.Rademaker JLW, Louws FJ, Schultz MH, Rossbach U, Vauterin L, Swings J, et al. A comprehensive species to strain taxonomic framework for Xanthomonas. Phytopathology. 2005;95:1098–1111. doi: 10.1094/PHYTO-95-1098. [DOI] [PubMed] [Google Scholar]
- 28.Karavina C, Mandumbu R, Parwada C, Tibugari H. A review of the occurrence, biology and management of common bacterial blight. J Agric Technol. 2011;7:1459–1474. [Google Scholar]
- 29.Audy P, Laroche A, Saindon G, Huang HC, Gilbertson RL. Detection of the bean common blight bacteria, Xanthomonas campestris pv. phaseoli and X.c.phaseoli var. fuscans, using the polymerase chain reaction. Phytopathology. 1994;84:1185–1192. doi: 10.1094/phyto-84-1185. [DOI] [Google Scholar]
- 30.Grimault V, Olivier V, Rolland M, Darrasse A, Jacques M-A. Detection of Xanthomonas axonopodis pv. phaseoli and Xanthomonas axonopodis pv. phaseoli var. fuscans on Phaseolus vulgaris (bean). International Rules Seed Testing Annexe to Chapter 7 : Seed Heal Testing Methods. 2014. pp. 7–021–71-20. [Google Scholar]
- 31.Bourguet D, Guillemaud T. The hidden and external costs of pesticide use. In: Lichtfouse E, editor. Sustainable Agriculture Reviews. 2016. [Google Scholar]
- 32.Dangl JL, Horvath DM, Staskawicz BJ. Pivoting the plant immune system from dissection to deployment. Science. 2013;341:746–751. doi: 10.1126/science.1236011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Alladassi BME, Nkalubo ST, Mukankusi C, Mwale ES, Gibson P, Edema R, et al. Inheritance of resistance to common bacterial blight in four selected common bean (Phaseolus vulgaris L.) genotypes. J Plant Breed Crop Sci. 2017;9:71–78. doi: 10.5897/JPBCS2017.0644. [DOI] [Google Scholar]
- 34.Opio AF, Allen DJ, Teri JM. Pathogenic variation in Xanthomonas campestris pv. phaseoli, the causal agent of common bacterial blight in Phaseolus beans. Plant Pathol. 1996;45:1126–1133. doi: 10.1046/j.1365-3059.1996.d01-187.x. [DOI] [Google Scholar]
- 35.Viteri DM, Singh SP. Response of 21 common beans of diverse origins to two strains of the common bacterial blight pathogen, Xanthomonas campestris pv. phaseoli. Euphytica. 2014;200:379–388. doi: 10.1007/s10681-014-1161-x. [DOI] [Google Scholar]
- 36.Miklas PN, Beaver JS, Grafton KF, Freytag GF. Registration of TARS VCI-4B multiple disease resistant dry bean Germplasm. Crop Sci. 1994;34:1415. doi: 10.2135/cropsci1994.0011183x003400050065x. [DOI] [Google Scholar]
- 37.Parker JPK. Interspecific transfer of common bacterial blight resistance from Phaseolus acutifolius a gray to Phaseolus vulgaris L. 1985. [Google Scholar]
- 38.Jung G, Skroch PW, Coyne DP, Nienhuis J, Arnaud-Santana E, Ariyarathne HM, et al. Molecular-marker-based genetic analysis of tepary bean-derived common bacterial blight resistance in different developmental stages of common bean. J Am Soc Horticult Sci. 1997;122:329–337. doi: 10.21273/jashs.122.3.329. [DOI] [Google Scholar]
- 39.Pedraza García F, Gallego GJ, Beebe SE, Tohme M. J. Marcadores SCAR y RAPD para la resitencia a la bacteriosis comun (CBB). In: Singh SP, Voysest O (eds) Taller de mejoramiento de frijol para el Siglo XXI: bases para una estrategia para America Latina. CIAT, Cali, CO. 1997;53:130–134. doi: 10.1017/CBO9781107415324.004.
- 40.Yu K, Park SJ, Poysa V. Marker-assisted selection of common beans for resistance to common bacterial blight: efficacy and economics. Plant Breed. 2000;119:411–415. doi: 10.1046/j.1439-0523.2000.00514.x. [DOI] [Google Scholar]
- 41.Yu K, Park SJ, Zhang B, Haffner M, Poysa V. An SSR marker in the nitrate reductase gene of common bean is tightly linked to a major gene conferring resistance to common bacterial blight. Euphytica. 2004;138:89–95. doi: 10.1023/B:EUPH.0000047077.75285.60. [DOI] [Google Scholar]
- 42.Park SO, Coyne DP, Mutlu N, Jung G, Steadman JR. Confirmation of molecular markers and flower color associated with QTL for resistance to common bacterial blight in common beans. J Am Soc Hortic Sci. 1999;124:519–526. doi: 10.21273/jashs.124.5.519. [DOI] [Google Scholar]
- 43.Miklas PN, Delorme R, Stone V, Daly MJ, Stavely JR, Steadman JR, et al. Bacterial, fungal, and viral disease resistance loci mapped in a recombinant inbred common bean population ('Dorado’/XAN 176) J Am Soc Hortic Sci. 2000;125:476–481. doi: 10.21273/jashs.125.4.476. [DOI] [Google Scholar]
- 44.Miklas PN, Coyne DP, Grafton KF, Mutlu N, Reiser J, Lindgren DT, et al. A major QTL for common bacterial blight resistance derives from the common bean great northern landrace cultivar Montana no. 5. Euphytica. 2003;131:137–146. doi: 10.1023/A:1023064814531. [DOI] [Google Scholar]
- 45.Kelly JD, Gepts P, Miklas PN, Coyne DP. Tagging and mapping of genes and QTL and molecular marker-assisted selection for traits of economic importance in bean and cowpea. F Crop Res. 2003;82:135–154. doi: 10.1016/S0378-4290(03)00034-0. [DOI] [Google Scholar]
- 46.Miklas PN, Kelly JD, Beebe SE, Blair MW. Common bean breeding for resistance against biotic and abiotic stresses: from classical to MAS breeding. Euphytica. 2006;147:105–131. doi: 10.1007/s10681-006-4600-5. [DOI] [Google Scholar]
- 47.Shi C, Chaudhary S, Yu K, Park SJ, Navabi A, McClean PE. Identification of candidate genes associated with CBB resistance in common bean HR45 (Phaseolus vulgaris L.) using cDNA-AFLP. Mol Biol Rep. 2011;38:75–81. doi: 10.1007/s11033-010-0079-1. [DOI] [PubMed] [Google Scholar]
- 48.Cooper DM. Identification and characterization of common bacterial blight resistance genes in the resistant common bean (Phaseolus vulgaris) variety OAC Rex. 2015. [Google Scholar]
- 49.Wu J, Zhu J, Wang L, Wang S. Genome-wide association study identifies NBS-LRR-encoding genes related with anthracnose and common bacterial blight in the common bean. Front Plant Sci. 2017;8:1398. doi: 10.3389/fpls.2017.01398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Nodari RO, Tsai SM, Guzmán P, Gilbertson RL, Gepts P. Toward an integrated linkage map of common bean. III mapping genetic factors controlling host-bacteria interactions. Genetics. 1993;134:341–350. doi: 10.1093/genetics/134.1.341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Darsonval A, Darrasse A, Durand K, Bureau C, Cesbron S, Jacques M-A. Adhesion and fitness in the bean Phyllosphere and transmission to seed of Xanthomonas fuscans subsp. fuscans. Mol Plant-Microbe Interact. 2009;22:747–757. doi: 10.1094/mpmi-22-6-0747. [DOI] [PubMed] [Google Scholar]
- 52.Rousseau C, Hunault G, Gaillard S, Bourbeillon J, Montiel G, Simier P, et al. Phenoplant: a web resource for the exploration of large chlorophyll fluorescence image datasets. Plant Methods. 2015;11:24. doi: 10.1186/s13007-015-0068-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Vlasova A, Capella-Gutiérrez S, Rendón-Anaya M, Hernández-Oñate M, Minoche AE, Erb I, et al. Genome and transcriptome analysis of the Mesoamerican common bean and the role of gene duplications in establishing tissue and temporal specialization of genes. Genome Biol. 2016;17:32. doi: 10.1186/s13059-016-0883-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Schwartz AR, Morbitzer R, Lahaye T, Staskawicz BJ. TALE-induced bHLH transcription factors that activate a pectate lyase contribute to water soaking in bacterial spot of tomato. Proc Natl Acad Sci. 2017;114:E897–E903. doi: 10.1073/pnas.1620407114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wilkins KE, Booher NJ, Wang L, Bogdanove AJ. TAL effectors and activation of predicted host targets distinguish Asian from African strains of the rice pathogen Xanthomonas oryzae pv oryzicola while strict conservation suggests universal importance of five TAL effectors. Front Plant Sci. 2015;6:536. doi: 10.3389/fpls.2015.00536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Cernadas RA, Camillo LR, Benedetti CE. Transcriptional analysis of the sweet orange interaction with the citrus canker pathogens Xanthomonas axonopodis pv. citri and Xanthomonas axonopodis pv. aurantifolii. Mol Plant Pathol. 2008;9:609–631. doi: 10.1111/j.1364-3703.2008.00486.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hu Y, Zhang J, Jia H, Sosso D, Li T, Frommer WB, et al. Lateral organ boundaries 1 is a disease susceptibility gene for citrus bacterial canker disease. Proc Natl Acad Sci. 2014;111:E521–E529. doi: 10.1073/pnas.1313271111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Supek F, Bošnjak M, Škunca N, Šmuc T. Revigo summarizes and visualizes long lists of gene ontology terms. PLoS One. 2011;6:e21800. doi: 10.1371/journal.pone.0021800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Tian T, Liu Y, Yan H, You Q, Yi X, Du Z, et al. AgriGO v2.0: a GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017;45:W122–W129. doi: 10.1093/nar/gkx382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mindrebo J, Nartey C, Seto Y, Burkart M, Noel J. Unveilling the functional diversity of the alpha-beta hydrolase fold in plants. Curr Opin Struct Biol. 2016;41:233–246. doi: 10.1016/j.sbi.2016.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Canonne J, Froidure-Nicolas S, Rivas S. Phospholipases in action during plant defense signaling. Plant Signal Behav. 2011;6:13–18. doi: 10.4161/psb.6.1.14037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kachroo P, Shanklin J, Shah J, Whittle EJ, Klessig DF. A fatty acid desaturase modulates the activation of defense signaling pathways in plants. Proc Natl Acad Sci. 2001;98:9448–9453. doi: 10.1073/pnas.151258398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Davies PJ. Plant hormones: biosynthesis, signal transduction, action!. Springer, Dordrecht. Copyright Information: Springer Science+Business Media B.V. 2010. 10.1007/978-1-4020-2686-7.
- 64.Los DA, Mironov KS, Allakhverdiev SI. Regulatory role of membrane fluidity in gene expression and physiological functions. Photosynth Res. 2013;116:489–509. doi: 10.1007/s11120-013-9823-4. [DOI] [PubMed] [Google Scholar]
- 65.Van Loon LC, Van Strien EA. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol Mol Plant Pathol. 1999;55:85–97. doi: 10.1006/pmpp.1999.0213. [DOI] [Google Scholar]
- 66.Argueso CT, Ferreira FJ, Epple P, To JPC, Hutchison CE, Schaller GE, et al. Two-component elements mediate interactions between cytokinin and salicylic acid in plant immunity. PLoS Genet. 2012;8. 10.1371/journal.pgen.1002448. [DOI] [PMC free article] [PubMed]
- 67.Hackbusch J, Richter K, Müller J, Salamini F, Uhrig JF. A central role of Arabidopsis thaliana ovate family proteins in networking and subcellular localization of 3-aa loop extension homeodomain proteins. Proc Natl Acad Sci. 2005;102:4908–4912. doi: 10.1073/pnas.0501181102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Wang S, Chang Y, Guo J, Zeng Q, Ellis BE, Chen JG. Arabidopsis ovate family proteins, a novel transcriptional repressor family, control multiple aspects of plant growth and development. PLoS One. 2011;6:e23896. doi: 10.1371/journal.pone.0023896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Wang S, Chang Y, Ellis B. Overview of OVATE FAMILY PROTEINS, a novel class of plant-specific growth regulators. Front Plant Sci. 2016;7:417. doi: 10.3389/fpls.2016.00417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Blair MW, Cortés AJ, Farmer AD, Huang W, Ambachew D, Varma Penmetsa R, et al. Uneven recombination rate and linkage disequilibrium across a reference SNP map for common bean (Phaseolus vulgaris L.) PLoS One. 2018;13:e0189597. doi: 10.1371/journal.pone.0189597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Chen NWG, Sévignac M, Thareau V, Magdelenat G, David P, Ashfield T, et al. Specific resistances against Pseudomonas syringae effectors AvrB and AvrRpm1 have evolved differently in common bean (Phaseolus vulgaris), soybean (Glycine max), and Arabidopsis thaliana. New Phytol. 2010;187:941–956. doi: 10.1111/j.1469-8137.2010.03337.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Freyre R, Skroch PW, Geffroy V, Adam-Blondon AF, Shirmohamadali A, Johnson WC, et al. Towards an integrated linkage map of common bean. 4. Development of a core linkage map and alignment of RFLP maps. Theor Appl Genet. 1998;97:847–856. doi: 10.1007/s001220050964. [DOI] [Google Scholar]
- 73.Geffroy V, Sévignac M, De Oliveira JCF, Fouilloux G, Skroch P, Thoquet P, et al. Inheritance of partial resistance against Colletotrichum lindemuthianum in Phaseolus vulgaris and co-localization of quantitative trait loci with genes involved in specific resistance. Mol Plant-Microbe Interact. 2000;13:287–296. doi: 10.1094/MPMI.2000.13.3.287. [DOI] [PubMed] [Google Scholar]
- 74.Geffroy V, Sévignac M, Billant P, Dron M, Langin T. Resistance to Colletotrichum lindemuthianum in Phaseolus vulgaris: a case study for mapping two independent genes. Theor Appl Genet. 2008;116:407–415. doi: 10.1007/s00122-007-0678-y. [DOI] [PubMed] [Google Scholar]
- 75.Grisi MCM, Blair MW, Gepts P, Brondani C, Pereira PAA, Brondani RPV. Genetic mapping of a new set of microsatellite markers in a reference common bean (Phaseolus vulgaris) population BAT93 x Jalo EEP558. Genet Mol Res. 2007;6:691–706. [PubMed] [Google Scholar]
- 76.Heilig JA, Beaver JS, Wright EM, Song Q, Kelly JD. QTL analysis of symbiotic nitrogen fixation in a black bean population. Crop Sci. 2017;57:118–129. doi: 10.2135/cropsci2016.05.0348. [DOI] [Google Scholar]
- 77.McClean PE, Lee RK, Otto C, Gepts P, Bassett MJ. Molecular and phenotypic mapping of genes controlling seed coat pattern and color in common bean (Phaseolus vulgaris L.) J Hered. 2002;39:148–152. doi: 10.1093/jhered/93.2.148. [DOI] [PubMed] [Google Scholar]
- 78.Nodari RO, Koinange EMK, Kelly JD, Gepts P. Towards an integrated linkage map of common bean - 1. Development of genomic DNA probes and levels of restriction fragment length polymorphism. Theor Appl Genet. 1992;84:186–192. doi: 10.1007/BF00223999. [DOI] [PubMed] [Google Scholar]
- 79.Ballester A-R, Norelli J, Burchard E, Abdelfattah A, Levin E, González-Candelas L, et al. Transcriptomic response of resistant (PI613981–Malus sieversii) and susceptible (“Royal Gala”) genotypes of apple to blue Mold (Penicillium expansum) infection. Front Plant Sci. 2017;8:1981. doi: 10.3389/fpls.2017.01981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Jain S, Chittem K, Brueggeman R, Osorno JM, Richards J, Nelson BD, et al. Comparative transcriptome analysis of resistant and susceptible common bean genotypes in response to soybean cyst nematode infection. PLoS One. 2016;11:e0159338. doi: 10.1371/journal.pone.0159338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Padder BA, Kamfwa K, Awale HE, Kelly JD. Transcriptome profiling of the Phaseolus vulgaris - Colletotrichum lindemuthianum Pathosystem. PLoS One. 2016;11:e0165823. doi: 10.1371/journal.pone.0165823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Rubio M, Ballester AR, Olivares PM, Castro de Moura M, Dicenta F, Martínez-Gómez P. Gene expression analysis of plum pox virus (Sharka) susceptibility/resistance in apricot (Prunus armeniaca L.) PLoS One. 2015;10:e0144670. doi: 10.1371/journal.pone.0144670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zeng W, Sun Z, Cai Z, Chen H, Lai Z, Yang S, et al. Comparative transcriptome analysis of soybean response to bean pyralid larvae. BMC Genomics. 2017;18:871. doi: 10.1186/s12864-017-4256-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Tsai TM, Chen YR, Kao TW, Tsay WS, Wu CP, Huang DD, et al. PaCDPK1, a gene encoding calcium-dependent protein kinase from orchid, Phalaenopsis amabilis, is induced by cold, wounding, and pathogen challenge. Plant Cell Rep. 2007;26:1899–1908. doi: 10.1007/s00299-007-0389-5. [DOI] [PubMed] [Google Scholar]
- 85.Murillo I, Jaeck E, Cordero MJ, San SB. Transcriptional activation of a maize calcium-dependent protein kinase gene in response to fungal elicitors and infection. Plant Mol Biol. 2001;45:145–158. doi: 10.1023/A:1006430707075. [DOI] [PubMed] [Google Scholar]
- 86.Lecourieux D, Raneva R, Pugin A. Calcium in plant defence-signalling pathways. New Phytol. 2006;171:249–269. doi: 10.1111/j.1469-8137.2006.01777.x. [DOI] [PubMed] [Google Scholar]
- 87.Seybold H, Trempel F, Ranf S, Scheel D, Romeis T, Lee J. Ca2+ signalling in plant immune response: from pattern recognition receptors to Ca2+ decoding mechanisms. New Phytol. 2014;204:782–790. doi: 10.1111/nph.13031. [DOI] [PubMed] [Google Scholar]
- 88.Büttner D, Bonas U. Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol Rev. 2009;34:107–133. doi: 10.1111/j.1574-6976.2009.00192.x. [DOI] [PubMed] [Google Scholar]
- 89.Bürger M, Chory J. Stressed out about hormones: how plants orchestrate immunity. Cell Host Microbe. 2019;26:163–172. doi: 10.1016/j.chom.2019.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Ben-David A, Bashan Y, Okon Y. Ethylene production in pepper (Capsicum annuum) leaves infected with Xanthomonas campestris pv. vesicatoria. Physiol Mol Plant Pathol. 1986;29:305–316. doi: 10.1016/S0048-4059(86)80047-9. [DOI] [Google Scholar]
- 91.Dutta S, Biggs RH, Biggs RH. Regulation of ethylene biosynthesis in citrus leaves infected with Xanthomonas campestris pv. citri. Physiol Plant. 1991:225–31. 10.1111/j.1399-3054.1991.tb00085.x.
- 92.Lund ST, Stall RE, Klee HJ. Ethylene regulates the susceptible response to pathogen infection in tomato. Am Soc Plant Physiol Ethyl. 1998:371–82. 10.1105/tpc.10.3.371. [DOI] [PMC free article] [PubMed]
- 93.Gervasi F, Ferrante P, Dettori MT, Scortichini M, Verde I. Transcriptome reprogramming of resistant and susceptible peach genotypes during Xanthomonas arboricola pv. pruni early leaf infection. PLoS One. 2018;13:1–21. doi: 10.1371/journal.pone.0196590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Zou J, Rodriguez-Zas S, Aldea M, Li M, Zhu J, Gonzalez DO, et al. Expression profiling soybean response to pseudomonas syringae reveals new defense-related genes and rapid HR-specific downregulation of photosynthesis. Mol Plant-Microbe Interact. 2005;18:1161–1174. doi: 10.1094/mpmi-18-1161. [DOI] [PubMed] [Google Scholar]
- 95.Lewis LA, Polanski K, de Torres-Zabala M, Jayaraman S, Bowden L, Moore J, et al. Transcriptional dynamics driving MAMP-triggered immunity and pathogen effector-mediated immunosuppression in Arabidopsis leaves following infection with Pseudomonas syringae pv tomato DC3000. Plant Cell. 2015;27:3038–3064. doi: 10.1105/tpc.15.00471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Lopez C, Soto M, Restrepo S, Piégu B, Cooke R, Delseny M, et al. Gene expression profile in response to Xanthomonas axonopodis pv. manihotis infection in cassava using a cDNA microarray. Plant Mol Biol. 2005;57:393–410. doi: 10.1007/s11103-004-7819-3. [DOI] [PubMed] [Google Scholar]
- 97.Bilgin DD, Zavala JA, Zhu J, Clough SJ, Ort DR, Delucia EH. Biotic stress globally downregulates photosynthesis genes. Plant Cell Environ. 2010;33:1597–1613. doi: 10.1111/j.1365-3040.2010.02167.x. [DOI] [PubMed] [Google Scholar]
- 98.Berger S, Sinha AK, Roitsch T. Plant physiology meets phytopathology: plant primary metabolism and plant-pathogen interactions. J Exp Bot. 2007;58:4019–4026. doi: 10.1093/jxb/erm298REVIEW. [DOI] [PubMed] [Google Scholar]
- 99.Bonfig KB, Schreiber U, Gabler A, Roitsch T, Berger S. Infection with virulent and avirulent P. syringae strains differentially affects photosynthesis and sink metabolism in Arabidopsis leaves. Planta. 2006;225:1–12. doi: 10.1007/s00425-006-0303-3. [DOI] [PubMed] [Google Scholar]
- 100.Scharte J, Schön H, Weis E. Photosynthesis and carbohydrate metabolism in tobacco leaves during an incompatible interaction with Phytophthora nicotianae. Plant Cell Environ. 2005;28:1421–1435. doi: 10.1111/j.1365-3040.2005.01380.x. [DOI] [Google Scholar]
- 101.Swarbrick PJ, Schulze-Lefert P, Scholes JD. Metabolic consequences of susceptibility and resistance (race-specific and broad-spectrum) in barley leaves challenged with powdery mildew. Plant Cell Environ. 2006;29:1061–1076. doi: 10.1111/j.1365-3040.2005.01472.x. [DOI] [PubMed] [Google Scholar]
- 102.Kato Y, Miura E, Ido K, Ifuku K, Sakamoto W. The variegated mutants lacking chloroplastic FtsHs are defective in D1 degradation and accumulate reactive oxygen species. Plant Physiol. 2009;151:1790–1801. doi: 10.1104/pp.109.146589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Garavaglia BS, Thomas L, Gottig N, Zimaro T, Garofalo CG, Gehring C, et al. Shedding light on the role of photosynthesis in pathogen colonization and host defense. Commun Integr Biol. 2010;3:382–384. doi: 10.4161/cib.3.4.12029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Choi HW, Klessig DF. DAMPs, MAMPs, and NAMPs in plant innate immunity. BMC Plant Biol. 2016;16:232. doi: 10.1186/s12870-016-0921-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Han Y, Zhu Q, Zhang Z, Meng K, Hou Y, Ban Q, et al. Analysis of xyloglucan endotransglycosylase/hydrolase (XTH) genes and diverse roles of isoenzymes during persimmon fruit development and postharvest softening. PLoS One. 2015;10:e0123668. doi: 10.1371/journal.pone.0123668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Cosgrove DJ. Plant expansins: diversity and interactions with plant cell walls. Curr Opin Plant Biol. 2015;25:162–172. doi: 10.1016/j.pbi.2015.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Cosgrove DJ. Growth of the plant cell wall. Nat Rev Mol Cell Biol. 2005;6:850–861. doi: 10.1038/nrm1746. [DOI] [PubMed] [Google Scholar]
- 108.Majer C, Hochholdinger F. Defining the boundaries: structure and function of LOB domain proteins. Trends Plant Sci. 2011;16:47–52. doi: 10.1016/j.tplants.2010.09.009. [DOI] [PubMed] [Google Scholar]
- 109.Grimplet J, Pimentel D, Agudelo-Romero P, Martinez-Zapater JM, Fortes AM. The LATERAL ORGAN BOUNDARIES domain gene family in grapevine: genome-wide characterization and expression analyses during developmental processes and stress responses. Sci Rep. 2017;7:15968. doi: 10.1038/s41598-017-16240-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Springer P. Transcription factors expressed in lateral organ boundaries: identification of downstream targets. Univ North Texas Libr UNT Digit Libr. 2010;10:1-11.
- 111.David P, Chen NWG, Pedrosa-Harand A, Thareau V, Sevignac M, Cannon SB, et al. A nomadic subtelomeric disease resistance gene cluster in common bean. Plant Physiol. 2009;151:1048–1065. doi: 10.1104/pp.109.142109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Meziadi C, Richard MMS, Derquennes A, Thareau V, Blanchet S, Gratias A, et al. Development of molecular markers linked to disease resistance genes in common bean based on whole genome sequence. Plant Sci. 2016;242:351–357. doi: 10.1016/j.plantsci.2015.09.006. [DOI] [PubMed] [Google Scholar]
- 113.Richard MMS, Thareau V, Chen NWG, Meziadi C, Pflieger S, Geffroy V, Pérez de la Vega M. In: What is present at common bean subtelomeres? Large resistance gene clusters, knobs and Khipu satellite DNA. Santalla M, Marsolais F, editors. 2017. [Google Scholar]
- 114.Chen NWG, Thareau V, Ribeiro T, Magdelenat G, Ashfield T, Innes RW, et al. Common bean subtelomeres are hot spots of recombination and favor resistance gene evolution. Front Plant Sci. 2018;9:1–15. doi: 10.3389/fpls.2018.01185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Boch J, Bonas U. Xanthomonas AvrBs3 family-type III effectors: discovery and function. Annu Rev Phytopathol. 2010;48:419–436. doi: 10.1146/annurev-phyto-080508-081936. [DOI] [PubMed] [Google Scholar]
- 116.Ruh M, Briand M, Bonneau S, Jacques M, Chen NWG. Xanthomonas adaptation to common bean is associated with horizontal transfers of genes encoding TAL effectors. BMC Genomics. 2017;18:1–18. doi: 10.1186/s12864-017-4087-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Boch J, Bonas U, Lahaye T. TAL effectors - pathogen strategies and plant resistance engineering. New Phytol. 2014;204:823–832. doi: 10.1111/nph.13015. [DOI] [PubMed] [Google Scholar]
- 118.Chen L. SWEET sugar transporters for phloem transport and pathogen nutrition. New Phytol. 2014;201:1150–1155. doi: 10.1111/nph.12445. [DOI] [PubMed] [Google Scholar]
- 119.Chen L, Hou B, Lalonde S, Takanaga H, Hartung ML, Qu X, et al. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature. 2010;468:527–532. doi: 10.1038/nature09606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Chen L, Qu X, Hou B, Sosso D, Osorio S, Fernie AR, et al. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science. 2012;335:207–211. doi: 10.1126/science.1213351. [DOI] [PubMed] [Google Scholar]
- 121.Zhou J, Peng Z, Long J, Sosso D, Liu B, Eom JS, et al. Gene targeting by the TAL effector PthXo2 reveals cryptic resistance gene for bacterial blight of rice. Plant J. 2015;82:632–643. doi: 10.1111/tpj.12838. [DOI] [PubMed] [Google Scholar]
- 122.Yu Y, Streubel J, Balzergue S, Champion A, Boch J, Koebnik R, et al. Colonization of rice leaf blades by an African strain of Xanthomonas oryzae pv. oryzae depends on a new TAL effector that induces the rice Nodulin−3 Os11N3 gene. Mol Plant-Microbe Interact. 2011;24:1102–1113. doi: 10.1094/MPMI-11-10-0254. [DOI] [PubMed] [Google Scholar]
- 123.Yang B, White FF. Diverse members of the AvrBs3/PthA family of type III effectors are major virulence determinants in bacterial blight disease of rice. Mol Plant-Microbe Interact. 2004;17:1192–1200. doi: 10.1094/MPMI.2004.17.11.1192. [DOI] [PubMed] [Google Scholar]
- 124.Yang B, Sugio A, White FF. Os8N3 is a host disease-susceptibility gene for bacterial blight of rice. Proc Natl Acad Sci. 2006;103:10503–10508. doi: 10.1073/pnas.0604088103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Antony G, Zhou J, Huang S, Li T, Liu B, White F, et al. Rice xa13 recessive resistance to bacterial blight is defeated by induction of the disease susceptibility gene Os-11N3. Plant Cell. 2010;22:3864–3876. doi: 10.1105/tpc.110.078964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Römer P, Recht S, Strauß T, Elsaesser J, Schornack S, Boch J, et al. Promoter elements of rice susceptibility genes are bound and activated by specific TAL effectors from the bacterial blight pathogen, Xanthomonas oryzae pv. oryzae. New Phytol. 2010;187:1048–1057. doi: 10.1111/j.1469-8137.2010.03217.x. [DOI] [PubMed] [Google Scholar]
- 127.Streubel J, Pesce C, Hutin M, Koebnik R, Boch J, Szurek B. Five phylogenetically close rice SWEET genes confer TAL effector-mediated susceptibility to Xanthomonas oryzae pv. oryzae. New Phytol. 2013;200:808–819. doi: 10.1111/nph.12411. [DOI] [PubMed] [Google Scholar]
- 128.Cohn M, Bart RS, Shybut M, Dahlbeck D, Gomez M, Morbitzer R, et al. Xanthomonas axonopodis virulence is promoted by a transcription activator-like effector-mediated induction of a SWEET sugar transporter in cassava. Mol Plant-Microbe Interact. 2014;27:1186–1198. doi: 10.1094/MPMI-06-14-0161-R. [DOI] [PubMed] [Google Scholar]
- 129.Cox KL, Meng F, Wilkins KE, Li F, Wang P, Booher NJ, et al. TAL effector driven induction of a SWEET gene confers susceptibility to bacterial blight of cotton. Nat Commun. 2017;24:15588. doi: 10.1038/ncomms15588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Tran TT, Pérez-Quintero AL, Wonni I, Carpenter SCD, Yu Y, Wang L, et al. Functional analysis of African Xanthomonas oryzae pv . oryzae TALomes reveals a new susceptibility gene in bacterial leaf light of rice. PLoS Pathog. 2018;14:e1007092. doi: 10.1371/journal.ppat.1007092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Kay S, Hahn S, Marois E, Hause G, Bonas U. A bacterial effector acts as a plant transcription factor and induces a cell size regulator. Science. 2007;318:648–651. doi: 10.1126/science.1144956. [DOI] [PubMed] [Google Scholar]
- 132.Al-Saadi A, Reddy JD, Duan YP, Brunings AM, Yuan Q, Gabriel DW. All five host-range variants of Xanthomonas citri carry one pthA homolog with 17.5 repeats that determines pathogenicity on citrus, but none determine host-range variation. Mol Plant-Microbe Interact. 2007;20:934–943. doi: 10.1094/MPMI-20-8-0934. [DOI] [PubMed] [Google Scholar]
- 133.Li Z, Zou L, Ye G, Xiong L, Ji Z, Zakria M, et al. A potential disease susceptibility gene CsLOB of citrus is targeted by a major virulence effector PthA of xanthomonas citri subsp. citri. Mol Plant. 2014;7:912–915. doi: 10.1093/mp/sst176. [DOI] [PubMed] [Google Scholar]
- 134.Rousseau C, Belin E, Bove E, Rousseau D, Fabre F, Berruyer R, et al. High throughput quantitative phenotyping of plant resistance using chlorophyll fluorescence image analysis. Plant Methods. 2013;9:17. doi: 10.1186/1746-4811-9-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Sallet E, Gouzy J, Schiex T. EuGene: An Automated Integrative Gene Finder for Eukaryotes and Prokaryotes. Methods Mol Biol. 2019;1962:97-120. 10.1007/978-1-4939-9173-0_6. [DOI] [PubMed]
- 136.Schmutz J, Cannon SB, Schlueter J, Ma J, Mitros T, Nelson W, et al. Genome sequence of the palaeopolyploid soybean. Nature. 2010;463:178–183. doi: 10.1038/ng.3008. [DOI] [PubMed] [Google Scholar]
- 137.Seppey M, Manni M, Zdobnov EM. BUSCO: assessing genome assembly and annotation completeness. Methods Mol Biol. 1962;2019:227–245. doi: 10.7551/mitpress/7458.003.0021. [DOI] [PubMed] [Google Scholar]
- 138.Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15:550. doi: 10.1186/s13059-014-0550-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Ser B (Methodological) 1995;57:289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x. [DOI] [Google Scholar]
- 140.Thimm O, Bläsing O, Gibon Y, Nagel A, Meyer S, Krüger P, et al. MAPMAN: a user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 2004;37:914–939. doi: 10.1111/j.1365-313X.2004.02016.x. [DOI] [PubMed] [Google Scholar]
- 141.Kanehisa M, Sato Y. KEGG mapper for inferring cellular functions from protein sequences. Protein Sci. 2020;29:28–35. doi: 10.1002/pro.3711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Zheng Y, Jiao C, Sun H, Rosli HG, Pombo MA, Zhang P, et al. iTAK: a program for genome-wide prediction and classification of plant transcription factors, transcriptional regulators, and protein kinases. Mol Plant. 2016;9:1667–1670. doi: 10.1016/j.molp.2016.09.014. [DOI] [PubMed] [Google Scholar]
- 143.Richard MMS, Gratias A, Thareau V, Do KK, Balzergue S, Joets J, et al. Genomic and epigenomic immunity in common bean: the unusual features of NB-LRR gene family. DNA Res. 2017;0:1–12. doi: 10.1093/dnares/dsx046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Untergasser A, Cutcutache I, Koressaar T, Ye J, Faircloth BC, Remm M, et al. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012;40:e115. doi: 10.1093/nar/gks596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3:research0034.1–researc0034.11. doi: 10.1186/gb-2002-3-7-research0034. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Figure S1 Statistical analysis of RNA-Seq data. Statistics were performed by using the total number of RNA-Seq reads mapped on the 20,787 homologs shared by BAT93 and JaloEEP558. Principal component analysis (A). Triangles and dots represent the barycenters of RNA-Seq data from BAT93 and JaloEEP558, respectively. Data from plants inoculated with H2O are in orange while data from plants inoculated with Xanthomonas strain CFBP6546R are in black. Pearson correlation matrix (B). Numbers represent the means of Pearson correlation coefficients calculated using three biological replicates per condition.
Additional file 2: Figure S2. Validation of RNA-Seq results by RT-qPCR analysis in BAT93 (A) and JaloEEP558 (B). Gene expression data were expressed according to the 2–ΔΔCt method (Vandesompele et al. 2002) [145], relatively to three housekeeping genes: Act11, EF1-α and IDE, and to the value of water inoculated plants.
Additional file 3 Figure S3 Numbers of DEGs in the different families of transcription factors. The list of common bean transcription factors was retrieved from the iTAK database (http://itak.feilab.net, Zheng et al. 2016) [142]. The numbers of induced and repressed genes are represented by red and blue bars respectively, in the specific transcriptomes of BAT93 (solid) or JaloEEP558 (hatched). Only families with at least 10 DEGs were represented.
Additional file 4: Figure S4. KEGG orthology map for plant hormone signal transduction. The KEGG orthology maps (pvu04075) of BAT93 (A) and JaloEEP558 (B) highlight DEGs that were induced (red), repressed (blue), or both induced and repressed (green) 48 h after inoculation with Xanthomonas phaseoli pv. phaseoli. The number of DEGs (colored numbers) and the average Log2FC (black numbers) are indicated above the boxes.
Additional file 5: Table S1. Number of genes shared among common bean genomes, as determined by reciprocal blastp (e-value <1E-6).
Additional file 6: Table S2. List of homologs shared by BAT93 and JaloEEP558.
Additional file 7: Table S3. List of DEGs in BAT93 and JaloEEP558 48 h after inoculation by X. phaseoli pv. phaseoli strain CFBP6546R.
Additional file 8: Table S4. Raw enrichment tests on the core, BAT93-specific or JaloEEP558-specific transcriptomes.
Additional file 9: Table S5. Enrichment tests on the core, BAT93-specific or JaloEEP558-specific transcriptomes, using induced or repressed DEGs separately.
Additional file 10: Table S6. List of enriched MapMan bins in the BAT93-specific and JaloEEP558-specific transcriptomes.
Additional file 12: Table S8. List of DEGs with |Δlog2FC| > 5.
Additional file 13: Table S9. List of DEGs with an inverse expression pattern between BAT93 and JaloEEP558.
Additional file 14: Table S10. List of DEGs in the four QRLs to CBB described in the progeny of BAT93xJaloEEP558 (Nodari et al., 1993) [50].
Additional file 15: Table S11. Genetic markers associated to the four QRLs to CBB described by Nodari et al. 1993 [the four QRLs to CBB described in the progeny of BAT93xJaloEEP558 (Nodari et al., 1993) [50].
Additional file 16: Table S12. Results of the enrichment tests according to the core, BAT93-specific or JaloEEP558-specific transcriptome and according to different log2FC threshold.
Additional file 17: Table S13. Primers used for RT-qPCR assays.
Data Availability Statement
Whole genome assemblies generated for this study have been submitted to GenBank under accession numbers JAAIFG000000000 (BAT93) and JAAIFH000000000 (JaloEEP58). Sequences and annotations are available at https://bbric-pipelines.toulouse.inra.fr/myGenomeBrowser?public=1. The raw RNA-Seq data and metadata have been submitted to the Gene Expression Omnibus under the accession number GSE155080 (SRA: SRP273448).