

Summary
CRISPR-based diagnostics (CRISPR-dx), including the Cas12-based DETECTR and Cas13-based SHERLOCK Class 2 CRISPRs, have been used to detect the presence of DNA or RNA from pathogens, such as the 2009 pandemic influenza virus A (IAV) and the 2019 novel coronavirus SARS-CoV-2. Here, we describe the collateral single-stranded DNA cleavage with Class 1 type I CRISPR-Cas3 and highlight its potential for development as a Cas3-mediated rapid (within 40 min), low-cost, instrument-free detection method for SARS-CoV-2. This assay, which we call Cas3-Operated Nucleic Acid detectioN (CONAN), not only detects SARS-CoV-2 in clinical samples, but also offers specific detection of single-base-pair mutations in IAV variants. This tool allows rapid and accurate point-of-care testing for patients with suspected SARS-CoV-2 or drug-resistant IAV infections in hospitals.
Subject areas: Diagnostics, Virology
Graphical abstract
Highlights
-
•
Type I CRISPR-Cas3 shows trans-cleavage activity on nearby nonspecific ssDNA
-
•
We developed CONAN as a method to easily and accurately detect viral RNA
-
•
CONAN provides rapid, low-cost, and instrument-free detection of SARS-CoV-2 and IAV
-
•
CONAN also allows the specific detection of single-base-pair mutations
Diagnostics; Virology
Introduction
Over the last 20 years the world has faced several epidemics and pandemics that have seriously threatened global public health, including the 2002 severe acute respiratory syndrome (SARS), the 2009 influenza virus A (IAV) pandemic, the 2012 Middle East respiratory syndrome (MERS), the 2014 Ebola outbreak, and the 2019 coronavirus disease (COVID-19) pandemic. This history highlights the urgent necessity for fast, sensitive, and specific diagnostic tools for virus surveillance, including antimicrobial resistant and emerging virus variants. Many countries use assays based on real-time reverse-transcriptase PCR (RT-qPCR) to detect viruses. However, the results of such assays on clinical samples from people with suspected virus infections are generally not ready until the day after sample collection because the samples need to be shipped to reference laboratories for accurate diagnostic testing (Bustin and Mueller, 2005; Corman et al., 2020; Ravina et al., 2020). RT-qPCR assays also require expensive equipment and well-trained personnel for their operation. Alternatively, rapid antigen tests can directly detect viral components without the amplification steps needed for RT-PCR; however, the evidence for their sensitivity and diagnostic accuracy require evaluation (Green and StGeorge, 2018). In contrast, serology antibody tests with lateral flow immunoassays can rapidly and sensitively determine the infection rate in a population, although IgG antibodies to viruses are generally only detectable 10–14 days postinfection (Kumar and Henrickson, 2012; Theel et al., 2020).
To overcome such limitations in the current diagnostic technologies, CRISPR-based diagnostics (CRISPR-dx) have been used to rapidly, robustly, and sensitively detect emerging viruses. These systems rely on type V CRISPR-Cas12, otherwise called DETECTR (DNA Endonuclease-TargEted CRISPR Trans Reporter) (Chen et al., 2018) or type VI CRISPR-Cas13, otherwise called SHERLOCK (Specific High-sensitive Enzymatic Reporter UnLOCKing) (Gootenberg et al., 2017). Both of these Cas enzymes, but not Cas9, exhibit nonspecific endonuclease activity in trans after binding to a specific cis target via programmable CRISPR RNAs (crRNAs). By combining isothermal amplification methods (e.g., Recombinase Polymerase Amplification, RPA (Piepenburg et al., 2006), or Loop-mediated isothermal AMPlification (LAMP) (Notomi et al., 2000) with reporting formats such as lateral flow detection with antigen-labeled reporters (Kellner et al., 2019), DETECTR (Broughton et al., 2020; Ding et al., 2020; Lucia et al., 2020) and SHERLOCK (Ackerman et al., 2020; Metsky et al., 2020) have recently been used for rapid and highly sensitive SARS-CoV-2 detection. Furthermore, during the peer reviewing period of this manuscript, type III CRISPR-Cas systems have also been reported for SARS-CoV-2 diagnosis (Santiago-Frangos et al., 2021). The Cas13-based strategy, PAC-MAN (Prophylactic Antiviral CRISPR in huMAN cells), has also been shown to inhibit and degrade SARS-CoV-2 viral RNA in respiratory epithelial cells (Abbott et al., 2020).
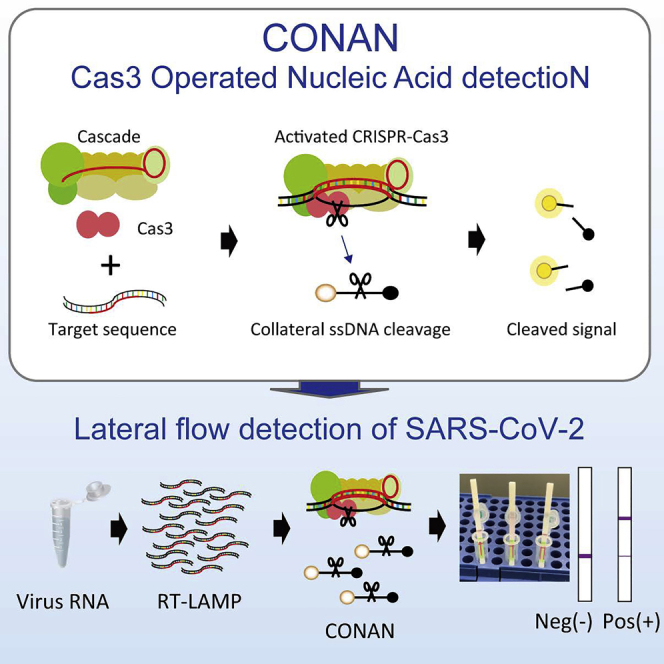
Class 2 CRISPRs (type V Cas12, type VI Cas13, and type II Cas9) only use a single Cas protein, whereas the three Class 1 CRISPRs (I, III, and IV) use multiple different Cas proteins. Several studies have reported on the mechanisms involved in type I CRISPR interference (Guo et al., 2017; Hochstrasser et al., 2014; Loeff et al., 2018; Mulepati and Bailey, 2013; Redding et al., 2015; Rutkauskas et al., 2015; van Erp et al., 2018; Westra et al., 2012; Xue et al., 2017). Type I CRISPR-Cas complexes have been reported to be seahorse-like structures containing Cas5, Cas6, multiple Cas7, Cas8 (Cse1), and two Cas11 (Cse2), and are named Cascade (Chowdhury et al., 2017; Hayes et al., 2016; Jackson et al., 2014; Mulepati et al., 2014; Sashital et al., 2012; Wiedenheft et al., 2011; Xiao et al., 2017). Cas3, a protein containing a serine-phenylalanine (SF) 2-helicase domain and a histidine-aspartic (HD) acid-nuclease domain, degrades its target DNA in a unidirectional ATP-dependent manner (Sinkunas et al., 2011; van Erp et al., 2018; Westra et al., 2012; Xiao et al., 2017, 2018). Interestingly, Streptococcus thermophiles Cas3 (Sinkunas et al., 2011) and Methanocaldococcus jannaschii Cas3 enzymes (Beloglazova et al., 2011) have an indiscriminate type of single-stranded DNA (ssDNA) cleavage when activated by Mg2+ bound to the catalytic site of the HD domain. In contrast, activation of the HD domain in Escherichia coli Cas3 (EcoCas3) (Mulepati and Bailey, 2013; Yoshimi et al., 2021) and Thermus thermophilus Cas3 (Mulepati and Bailey, 2011) is elicited by transition metal ions such as Co2+ and Ni2+, but not by Ca2+ and Mg2+. Despite type I-E E. coli CRISPR being one of the most thoroughly biochemically characterized in vitro plasmid DNA degradation-inducing systems (Hidalgo-Cantabrana and Barrangou, 2020; Zheng et al., 2020), whether the CRISPR-Cas3 system can mediate the target-activated, nonspecific ssDNA cleavage reported for Cas12 and Cas13 (Chen et al., 2018; Gootenberg et al., 2017) remains an open question. Here, we report on the third CRISPR-dx platform, Cas3-Operated Nucleic Acid detectioN (CONAN) (Figure 1A). When combined with isothermal amplification methods, CONAN provides a rapid, sensitive, and instrument-free detection system for SARS-CoV-2 point-of-care test (POCT) applications.
Figure 1.

Cas3-operated nucleic acid detection (CONAN)
(A) Schematic representation of the CONAN in vitro nucleic acid-detection platform. The E. coli CRISPR-Cas3 complex contains Cas3, Cas5, Cas6, Cas7, Cas8, and Cas11 proteins and CRISPR RNA (crRNA), FQ-ssDNA, fluorophore, and quencher-labeled single-stranded DNA probe.
(B) Collateral ssDNA cleavage activity measured by incubation of EcoCas3-EcoCascade/crRNA complex with a 60-bp dsDNA activator containing a target sequence flanked by a PAM and an FQ-labeled ssDNA probe in reaction buffer containing MgCl2, CoCl2, and ATP for 10 min at 37 °C. CRISPR-Cas3 mediated collateral ssDNA cleavage after targeting hEMX1-dsDNA in fragments with PAM (AAG, red), but not in fragments with non-PAM (CCA, blue), quantitatively represented by relative fluorescent units (RFU) per min; increasing rate of RFU/min (right). Means (n = 3), and standard deviations.
(C) CONAN assay on isothermal RPA amplicon products (blue) detected a single copy of the EMX1 activator fragments (1.7 a.m.); RFU at 10 min. Means (n = 3), and standard deviations.
(D) CONAN RPA also detected a single-copy mTyr activator (1.7 a.m.) when mixed with mouse genomic DNA. Means (n = 3), and standard deviations.
Results
Trans-cleavage activity of the type I CRISPR-Cas3 system
Owing to the poor solubility and easy aggregation of EcoCas3 protein at 37 °C (Beloglazova et al., 2011; Mulepati and Bailey, 2013; Sinkunas et al., 2011; Westra et al., 2012), we purified it using the baculovirus expression system (Hitchman et al., 2009) with Sf9 insect cells cultured at 20°C (Figure S1 and STAR Methods). EcoCas3 did not show nonspecific ssDNA cleavage activity in Mg2+-containing buffer, consistent with previous results (Mulepati and Bailey, 2013), whereas Thermobifida fusca Cas3 (TfuCas3) (Dolan et al., 2019) did show indiscriminate cleavage activity (Figure S2). We next co-expressed and size exclusion chromatographically (SEC)-purified the core complex of E. coli Cascade (EcoCascade) proteins and crRNA from E. coli JM109 at 37°C (Figure S1 and STAR Methods). To investigate whether the type I CRISPR-Cas3 system carries its trans-cleavage activity on nearby nonspecific ssDNAs in a similar fashion as type V Cas12 and type VI Cas13, we used the collateral cleavage assay previously reported for DETECTR (Chen et al., 2018) and SHERLOCK (Gootenberg et al., 2017). Briefly, we assembled the EcoCas3/EcoCascade-crRNA complex, a 60-bp double-stranded DNA (dsDNA) actftor for the specific target gene, human EMX1 (hEMX1) or mouse Tyr (mTyr) (Tables S1 and S2), and a fluorophore quencher (FQ)-labeled ssDNA probe in the Mg2+ reaction buffer (Chen et al., 2018; Gootenberg et al., 2017; Mulepati and Bailey, 2013). After 10 min incubation at 37 °C, we observed nonspecific trans-ssDNA cleavage from the EcoCas3/EcoCascade after target-specific dsDNA cleavage, which recognized the protospacer adjacent motif (PAM) (AAG), but not the non-PAM (CCA) (Figure 1B).
We next performed several additional experiments to understand the biochemical activities necessary for collateral cleavage activity by the Cascade-Cas3 system. First, the longer 3-Kb dsDNA activator (hEMX1 plasmid), instead of the 60-bp short fragments, similarly mediated the trans-ssDNA cleavage by EcoCas3/EcoCascade (Figure S3). Second, changes in the concentration of the FQ-ssDNA probe clearly correlated with the signal intensity (Figure S4), suggesting that Cas3 can cleave several copies of ssDNA per bound Cascade. Third, in ATP-free reaction buffer (−), the collateral activity of the EcoCas3 protein was similar to that of wild-type EcoCas3 and the SF2 motif III S483A/T485A (dead helicase mutant, dhCas3) mutant (Morisaka et al., 2019) in ATP (+) buffer, meaning that the helicase activity of EcoCas3 is not essential for collateral cleavage (Figure S5). Finally, omitting Mg2+ and Co2+ from the reaction buffer weakened this trans-ssDNA cleavage activity, whereas adding a divalent ion chelating agent (EDTA) abolished the activity (Figure S6). Therefore, we named this protocol Cas3-Operated Nucleic Acid detectioN (CONAN) (Figure 1A).
CONAN: in vitro nucleic acid-detection platform
We then investigated the detection sensitivity of CONAN by diluting the hEMX1 or mTyr dsDNA activator with Cas3, Cascade, and FQ-ssDNA in the reaction buffer. CONAN’|'s limit of detection (LoD) was >1.0 × 1010 copies for the activator (Figure S7). To improve the LoD, we performed isothermal RPA (TwistAmp Basic kit, Maidenhead, UK) at 37 °C for 30 min, followed by a 10-min incubation with Cascade and Cas3, thereby determining the activator’|'s single-copy sensitivity level (∼1.7 a.m.) (Figure 1C). We also achieved robust detection of the attomolar (aM) level activator by CONAN after mixing it with mouse genomic DNA (Figure 1D). Detecting FQ-ssDNA cleavage needs a microplate reader for fluorescence intensity measurement. Instead of this laboratory instrument, a lateral flow strip can be used for instrument-free and portable diagnosis by the virus POCT (Kellner et al., 2019; Lucia et al., 2020). In principle, abundant reporter accumulates anti-FITC antibody-gold nanoparticle conjugates at the first line (negative) on the strip, whereas cleavage of the reporter would reduce accumulation on the first line and result in signal on the second line (positive) with <2 min of flow (Figure S8). Using this lateral flow strip we performed a one-pot assay with CONAN-RPA, thereby detecting a single copy level of the target dsDNA within 1 hour (Figure 1D).
CONAN-based assay for rapid detection of SARS-CoV-2 and influenza virus
We next examined whether the CONAN lateral flow assay would be effective for SARS-CoV-2 diagnosis (Figure 2A), as has been recently reported for DETECTR (Broughton et al., 2020; Ding et al., 2020; Lucia et al., 2020) and for SHERLOCK (Ackerman et al., 2020; Metsky et al., 2020). We designed primers to amplify the N (nucleoprotein) gene regions (N1 and N2) from SARS-CoV-2 (Table S3), which overlap with the region used in the DETECTR-based assay (Broughton et al., 2020), along with primers for the RT-qPCR assay from the United States Centers for Disease Control (US CDC) (Kimball et al., 2020). The RT-qPCR assay successfully amplified both the N1 and N2 regions of SARS-CoV-2, with LoDs of <102 copies (Figure 2B). However, RT-RPA followed by Cas3-based CONAN or Cas12a-based DETECTR in the one-step 37 °C 30 min incubation (Table S1) did not effectively detect SARS-CoV-2, probably because the N1 and N2 primers we designed for RT-RPA (Table S3) did not match the sensitivity of the CRISPR-based assay (Figure S9).
Figure 2.

CRISPR-Cas3-based assay for rapid detection of SARS-CoV-2 and influenza virus
(A) Schematic representation of the CONAN SARS-CoV-2 detection assay including a conventional RNA extraction step, RT-LAMP (62 °C, 20–30 min), CONAN (37 °C, 10 min), and lateral flow (RT, 2 min).
(B) The limit of detection (LoD) of the US CDC’|'s RT-qPCR assay amplification of the N1 and N2 regions of the SARS-CoV-2 N gene. Cq, cycle quantification value.
(C) LoD of CONAN-based and DETECTR-based assays for the N1 and N2 region of SARS-CoV-2. RFU, relative fluorescence unit.
(D) LoD of the CONAN-based lateral flow assay for the N1 region of SARS-CoV-2. CONAN’|'s LoD was 2 min incubation (left) and <102 copies (right). Positive (red arrow) and negative (black arrow) bands for CONAN (see Figure S8).
(E) Comparison of SARS-CoV-2 CONAN and DETECTR assays on 31 clinical samples (10 positive and 21 negative for SARS-CoV-2 by the CDC RT-qPCR assay) (see also Figure S10).
(F) CONAN-based assay for detecting I38T variants in influenza viruses. Means (n = 3) and standard deviations. ∗p < 0.01, one-way ANOVA with post-hoc test.
(G) CONAN-based assay for detecting I222R, H274Y, and N294S variants in influenza viruses. Means (n = 3) and standard deviations. ∗p < 0.01, one-way ANOVA with post-hoc test.
Designing primers for LAMP assays can be complicated, but the upside is that these assays seem to be less sensitive to inhibitors or off-target nucleic acid contamination in the samples (Francois et al., 2011; Notomi et al., 2000). RT and LAMP at 62 °C for 30 min, followed by CONAN or DETECTR for 10 min at 37 °C, both specifically detected SARS-CoV-2 with crRNA–N1 and –N2 (Figure 2C). The LoDs for CONAN-LAMP and DETECTR-LAMP (<102 copies) compared favorably with the CDC’|'s RT-qPCR assay for SARS-CoV-2 detection (Broughton et al., 2020; Corman et al., 2020; Udugama et al., 2020). This rapid detection by CONAN-LAMP was achieved at 62 °C for 30 min, with the lateral flow strip achieving a LoD of <102 copies for SARS-CoV-2 (Figure 2D and Video S1).
We then tested the extracted RNA samples from 10 PCR-positive COVID-19 patients and 21 PCR-negative samples from nasopharyngeal swabs, using the CONAN RT-LAMP and DETCTR RT-LAMP assays with lateral flow strip readouts (Figures 2E and S10). SARS-CoV-2 was detected by the CONAN RT-LAMP assay in nine of 10 patient swabs and detected in one of 21 negative swab samples (positive predictive agreement, 90%; negative predictive agreement, 95%). One negative swab from a COVID-19 patient was confirmed to be below the established LoD of <102 copies (Figure S10). This 94% detection rate (29/31) is comparable with that of the DETECTR RT-LAMP assay in this study (Figure 2E) and that previously reported for a COVID-19 POCT (Broughton et al., 2020).
Finally, we investigated whether CONAN was able to discriminate a single-base-pair mutation in the target sequences, as previously shown by Cas13 SHERLOCK (Gootenberg et al., 2017, 2018) and by Cas12b CDetection (Teng et al., 2019). The I38T drug-resistance mutation in influenza virus A confers reduced susceptibility to baloxavir treatment for IAV infections (Imai et al., 2020; Sato et al., 2020). We designed a set of crRNAs for CONAN specifically targeting the I38T variant in IAV H1N1pdm09 and H3N2 strains (Table S1). After incubation, the CONAN assay clearly only detected wild-type (WT) H1N1pdm09 and H3N2 strains, but not I38T variants (Figure 2F). To further evaluate the specificity of single-base-pair discrimination by CONAN, several crRNAs were also designed to detect the neuraminidase mutations that reduce viral susceptibility to oseltamivir in the IAV H1N1pdm09 strain (I222R and H274Y) and in the IAV H3N2 strain (N294S) (Hatakeyama et al., 2007; McKimm-Breschkin, 2013) (Table S1). Again, the CONAN assay specifically identified all three mutations (encoding I222R, H274Y, and N294S), but not the corresponding WT positions (Figure 2G). Thus, the accurate SNP-level detection by CONAN should facilitate quick and mobile POCT in hospitals and other medical facilities.
Discussion
In this study, we reported on the discovery of Class I Cas3-mediated collateral trans cleavage activity. Our newly developed CONAN lateral flow assay, which uses this type of collateral cleavage, facilitated the rapid, robust, and sensitive detection of the novel coronavirus, SARS-CoV-2. Unlike the Class II single Cas12 and Cas13 platforms, CONAN employs multiple Cas proteins (Cas3, 5, 6, 7, 8, and 11), which can be premixed in the reaction buffer. By screening mismatches for each nucleotide in the 32-nt spacer, we found that a single mismatch in the spacer region, even within the seed region (positions 1–8), resulted in little or no effect on collateral ssDNA cleavage activity (Figure S11 and Table S4). This is supported by recently reported results showing that Cas12a has multiple nicking activities with tolerance of four to eight mismatches within the spacer sequences (Fu et al., 2019; Murugan et al., 2020). In contrast, CONAN shows high specificity for single-base-pair discrimination within the PAM site, supporting the applicability of CONAN-based detection assays for POCTs, even on novel emerging coronavirus mutants such as the A,B,C types (Forster et al., 2020) as well as spike protein mutations (Korber et al., 2020).
In summary, we have shown that CONAN, a Cas3-based novel in vitro nucleic acid-detection platform, is a rapid (within 30–40 min), low-cost, and instrument-free detection method for SARS-CoV-2. We also showed that this CONAN-based assay enables single-base-pair discrimination (Figures 2F and 2G) (Yoshimi et al., 2021), thereby facilitating the deployment of CRISPR-dx for quick and mobile POCTs for drug-resistant IAV variants in hospitals or clinics.
Limitations of the study
Although we have discovered the collateral ssDNA cleavage activity in the EcoCascade-Cas3 system, this study does not describe the mechanisms underlying how Cas3 mediates the collateral ssDNA cleavages as well as targeted double-stranded DNA cleavages. Our preprint manuscript describes several insights for these molecular mechanisms (Yoshimi et al., 2021).
STAR★Methods
Key resources table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Bacterial and virus strains | ||
| E. coli: MAX Efficiency™ DH5α Competent Cells | Thermo Fisher Scientific | Cat#18258012 |
| E. coli: MAX Efficiency™ DH10Bac Competent Cells | Thermo Fisher Scientific | Cat#10361012 |
| E. coli: JM109(DE3) | Promega | Cat#P980A |
| Biological samples | ||
| SARS-CoV-2 clinical samples | IMSUT hospital | N/A |
| E.coli Cascade-crRNA complex, see Table S1 | In-house | N/A |
| E.coli Cas3 protein | In-house | N/A |
| LbCas12a (Cpf1) protein | Integrated DNA Technologies | Cat#10007922 |
| Chemicals, peptides, and recombinant proteins | ||
| ATP | TaKaRa | Cat#4041 |
| BpiI (BbsI) | Thermo Fisher Scientific | Cat#ER1011 |
| DNA ligation mix | TaKaRa | Cat#6023 |
| Tks Gflex™ DNA polymerase | TaKaRa | Cat#R060A |
| ExoSAP-IT™ Express PCR product cleanup reagent | Applied Biosystems | Cat#75001.1.ML |
| HEPES-KOH pH 7.5 | NACALAI TESQUE | Cat#15639-84 |
| Ni-NTA agarose | Qiagen | Cat#30210 |
| Superdex 200 Increase 10/300 GL | Cytiva | Cat#28990944 |
| PSFM-J1 medium | Fujifilm-Wako | Cat#160-25851 |
| FBS | Thermo Fisher Scientific | Cat#10270-106 |
| 2-YT Broth | Thermo Fisher Scientific | Cat #22712020 |
| IPTG (Isopropyl-β-D-thiogalactopyranoside) | TaKaRa | Cat#9030 |
| FuGENE® HD transfection reagent | Promega | Cat#E2311 |
| Critical commercial assays | ||
| PureLink HiPure plasmid filter midiprep kit | Thermo Fisher Scientific | Cat#K210015 |
| TwistAmp Basic | TwistDX | Cat#TABAS03KIT |
| WarmStart LAMP kit (DNA & RNA) | NEB | Cat#E1700S |
| HybriDetect – universal lateral flow assay kit | Milenia Biotec | Cat#MGHD 1 |
| SARS-CoV-2 direct detection RT-qPCR kit | TaKaRa | Cat#RC300A |
| Bac-to-Bac™ vector kit | Thermo Fisher Scientific | Cat#10360014 |
| Experimental models: cell lines | ||
| VeroE6/TMPRSS2 | Japanese Collection of Research Bioresources Cell Bank |
https://cellbank.nibiohn.go.jp/english/ no. JCRB1819 |
| Sf9 cells | Thermo Fisher Scientific | Cat#11496015 |
| Oligonucleotides | ||
| Primers, see Table S3 | Eurofins Genomics | N/A |
| Primers with quenchers and probes, see Table S3 | Integrated DNA Technologies | N/A |
| Recombinant DNA | ||
| pRSFDuet-1 | Merck Millipore | Cat#71341-3CN |
| pCDFDuet-1 | Merck Millipore | Cat#71340-3CN |
| pACYCDuet-1 | Merck Millipore | Cat#71147-3CN |
| pACYCDuet-1_crRNA-empty-Bbs1v2 | In-house | N/A |
| pCDFDuet-1_6His-Cas11 | In-house | N/A |
| pRSFDuet-1_ Cas5-6-7-8-11 | In-house | N/A |
| pFastbac-1_8HisCas3 | In-house | N/A |
| Software and algorithms | ||
| ImageJ | (Schneider et al., 2012) | https://imagej.nih.gov/ij/ |
| Prism 8 | GraphPad | https://www.graphpad.com/scientific-software/prism |
| PrimerExplorer v.5 | Eiken Chemical | https://primerexplorer.jp/ |
| Other | ||
| Thermal cycler | Bio-rad | Cat#T100 |
| qPCR instruments | Bio-rad | Cat#CFX Connect |
| New Brunswick™ Innova® 44R bioshaker | Eppendorf | Cat#M1282-0007 |
| BR-43FL MR bioshaker | TAITEC | Cat#0053027-000 |
| Cool Incubator i-CUBE | AS ONE | Cat#FCI-280HG |
| AKTA pure 25 | Cytiva | Cat#29018225 |
Resource availability
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the Lead Contact, Tomoji Mashimo (mashimo@ims.u-tokyo.sc.jp).
Materials availability
All reagents and materials used in this manuscript are available upon request or prepared to be available from commercial sources.
Experimental model and subject details
TMPRSS2-expressing VeroE6 cells and Sf9 cells
Vero E6 cells expressing human serine protease TMPRSS2 (VeroE6-TMPRSS2), derived from the kidney of an African green monkey, were kindly provided from National Institute of Infectious Diseases. The cells are available from the Japanese Collection of Research Bioresources Cell Bank in Japan (https://cellbank.nibiohn.go.jp/english/) (JCRB no. JCRB1819). The VeroE6-TMPRSS2 cells were incubated at 37°C under 5% CO2 in Dulbecco's Modified Eagle Medium (DMEM) containing 10% fetal calf serum (FCS), 1 mg/ml G418, 100 units/ml penicillin, 100 μg/ml streptomycin, and 5 μg/ml plasmocin prophylactic (InvivoGen, CA, USA). Sf9 cells were obtained from Thermo Fisher Scientific (Waltham, Massachusetts, USA) and maintained at 28°C with 130 rpm in PSFM-J1 medium (Fujifilm-wako, Osaka, Japan).
Baculovirus
Baculoviruses for EcoCas3 protein were produced using the Bac-to-Bac™ vector kit (Thermo Fisher Scientific) following to the manufacturer’s protocol. Briefly, EcoCas3 cDNA were cloned into the pFastbac-1 plasmids, which were transformed to the MAX Efficiency™ DH10Bac Competent cells (Thermo Fisher Scientific) in LB medium. After the collection of the recombinant bacmid DNA by single colony screening, the bacmid DNA were transfected to the Sf9 cells by FuGENE HD transfection reagent in PSFM-J1 medium. We incubated the cells at 27 °C for 72 hours and collected the supernatant as a first virus stock after the centrifuge of 3,500 rpm for 5 minutes. To increase the virus titer, 15 ml of the virus stock were added to the Sf9 cells at 0.6 × 106 cells/ml in 100 ml of PSFM-J1 medium. We again collected the supernatant as a baculovirus stock and used it for further experiments.
Collection of human clinical samples
Clinical nasopharyngeal and oropharyngeal swab samples from patients infected with SARS-CoV-2 were collected by IMSUT hospital (The University of Tokyo). Negative nasopharyngeal swabs were collected from healthy donors at IMSUT. RNA from the samples of patients and healthy donors was extracted as described in the NIID-approved protocol (input 140 μl; elution, 60 μl) using the Viral RNA mini kit (QIAGEN). These clinical specimens were discriminated by RT-PCR for positive or negative for SARS-CoV-2 infection prior to CRISPR-based assays. We used small sample size (10 positive and 21 negative) to validate the CONAN-based assays in this study, but more trials with a larger sample size will be needed before considering for clinical application.
The information about the patients and healthy individuals such as the age and gender are not available due to ethical and privacy restriction. However, we don’t anticipate the sex or age of the participants to have any influence on the results. Human samples were collected by following protocols approved by the Research Ethics Review Committee of the Institute of Medical Science, the University of Tokyo (approval number 2019–71–0201).
Methods details
CRISPR preparation
Cascade/Cas3 and Cas12a target sites were based on the human EMX1 gene, the mouse Tyr gene, and the N gene from SARS-CoV-2 (Table S1). A baculovirus expression system was used to purify the EcoCas3 protein, as previously shown (Yoshimi et al., 2021). Briefly, EcoCas3 cDNA was cloned using an octa-histidine tag and a six asparagine-histidine repeat tag into pFastbac-1 plasmids. Sf9 cells were infected with baculovirus at a multiplicity of infection (MOI) of two with 2% FBS in PSFM-J1 medium at 28°C with 130 rpm for 24 h. We changed the culture temperature to 20°C with 110 rpm for 4 d. The expressed EcoCas3 protein was purified using nickel affinity resin (Ni-NTA, QIAGEN, Venlo, the Netherlands). To remove the tags, purified EcoCas3 was digested with TEV protease, and further purified by size-exclusion chromatography (Superdex 200 Increase 10/300 GL; Thermo Fisher Scientific) in 0.2 M NaCl, 10% glycerol, 1 mM DTT, and 20 mM HEPES-Na (pH 7.0). Purified EcoCas3 protein was evaluated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (Figure S1A).
The EcoCascade and crRNA complex were purified in accordance with previously reported methods (Hochstrasser et al., 2014; Jore et al., 2011; Yoshimi et al., 2021). Briefly, EcoCascade/crRNA ribonucleoproteins (RNPs) were expressed in JM109 (DE3) by co-transformation with three plasmids: one plasmid encoding a hexahistidine tag and HRV3C protease recognition site in the N-terminus of Cas11 (plasmid pCDFDuet-1); one plasmid containing the EcoCascade operon and the genes encoding Cas5, Cas6, Cas7, Cas8, and Cas11 (plasmid pRSFDuet-1); and the final plasmid encoding crRNA (pACYCDuet-1) (Yoshimi et al., 2021). The transformed bacteria were cultured in 2xYT medium at 37 °C with 130 rpm. After the OD600 became 0.6 to 0.8, we added IPTG (final concentration 0.4 mM) and cultured at 26 °C with 110 rpm for 16 hours. The expressed Cascade-crRNA RNPs were purified by Ni-NTA resin. After removing the hexahistidine tag using HRV3C protease, the EcoCascade/crRNA RNPs were further purified by size-exclusion chromatography in 350 mM NaCl, 1 mM DTT, and 20 mM HEPES-Na (pH 7.0) and size-evaluated by SDS-PAGE (Figure S1B). RNP sizes were found to be consistent with those of previous reports (Hochstrasser et al., 2014; Jore et al., 2011). LbCas12a were purchased from Integrated DNA Technologies (IDT, Coralville, IA). Target-specific crRNAs were also purchased from IDT.
DNA and RNA preparation
For the Cascade/Cas3 activator templates, DNA fragments of hEMX1, mTyr and IAV variants (which include a target site) were designed and purchased from IDT (Table S2). Total mouse genomic DNA from a C57BL/6 strain was used after purification (Maxwell RSC Cell DNA Purification Kit; Promega, Madison, Wisconsin). LAMP SARS-CoV-2 primers were designed against regions of the N gene using PrimerExplorer v.5 (Eiken Chemical Co.; https://primerexplorer.jp/). The primers used for isothermal PCR are listed in Table S3.
Viral RNAs from SARS-CoV-2 were prepared according to the established protocol from the National Institute of Infectious Diseases in Japan (Matsuyama et al., 2020). Viral RNAs were purified from an infected TMPRSS2-expressing VeroE6 cell line using the QIAamp Viral RNA Mini Kit (QIAGEN) according to the manufacturer’s protocol.
Real-time RT-PCR
RT-qPCR was used to determine SARS-CoV-2 RNA copy numbers using Reliance One-Step Multiplex RT-qPCR Supermix and CFX Connect (Bio-Rad Laboratories, Hercules, CA) according to the manufacturer’s protocols. N gene-specific primer and probe sets for RT-qPCR assays and the SARS-CoV-2 plasmid (positive control) were purchased from IDT (Table S3). Copy numbers of SARS-CoV-2 were determined based on the regression line (y = -3.3125x + 40.527), which was generated using the serial-diluted plasmids and the N2-primer set.
CONAN assay
To characterize the Cas3 collateral cleavage assays, DNA templates were added to 100 nM Cascade-crRNA complex, 250 nM Cas3 and 2.5 mM ATP in CRISPR-Cas3 system working buffer (60 mM KCl, 10 mM MgCl2, 10 μM CoCl2, 5 mM HEPES-KOH pH 7.5), as previously described (Yoshimi et al., 2021). The ssDNA reporter probe (5ʹ-/5HEX/AAGGTCGGA/ZEN/GTCAACGGATTTGGTC/3IBFQ/-3ʹ) (250 nM) was added, and the probe’s cleavage-related change in the fluorescence signal was measured every 30 s for 10 min under 37 °C incubation.
To detect DNAs, isothermal amplification and RPA were performed using the TwistAmp Basic kit (TwistDx, Maidenhead, UK) according to the manufacturer’s protocol. Template DNAs were amplified by incubation at 37 °C for 20 min. To detect RNAs, isothermal amplification by the RT-LAMP method was performed using the WarmStart LAMP kit (NEB) according to the manufacturer’s protocol. Template RNAs were reverse transcribed and amplified by incubation at 62 °C for 20 min. To detect low-copy-number molecules in the patients’ samples, the incubation time was extended to 45 min.
The CRISPR-Cas3 system reaction mixture used for the CONAN method contained 100 nM EcoCascade-crRNA complex, 400 nM Cas3 and 2.5 mM ATP in the working buffer. LbCas12a were prepared as described previously12 for use as positive controls for trans cleavage activity. Cas12a (50 nM) was incubated with 62.5 nM of crRNA in 1× NEBuffer 2.1 for 30 min at 37 °C. After amplification, 2 μl of the amplicon was combined with 18 μl of Cas3 and the Cascade-crRNA complex or the Cas12a-crRNA complex, and 250 nM of the ssDNA reporter probe was added. The fluorescence signal was measured every 30 s at 37 °C.
Lateral flow assay
To optimize the CONAN assay for lateral flow readouts, the CRISPR-Cas3 system reaction mixture was added to 200 nM of the Cascade-crRNA complex, with 400 nM Cas3 and 2.5 mM ATP in the working buffer. A 2 μl aliquot of the amplicon was added to 18 μl Cas3, Cascade-crRNA complex and 500 nM ssDNA reporter probe (5ʹ-/5-FITC/TAGCATGTCA/3-Biotin/-3ʹ). The mixture was incubated for 10 min at 37 °C. After adding 50 μl nuclease-free water, a lateral flow strip (Milenia HybriDetect 1; TwistDx) was added to the reaction tube and the result was visualized after approximately 2 min. A lower band close to the sample pad indicated a negative result (uncut probes), whereas an upper band close to the top of the strip indicated the 5′ end of the cut probes. Emergence of an upper band indicated a positive result (Figure S8). The test band intensities on the lateral flow strips were quantified by the gray values using the ImageJ tool and were visualized on a heat map (Figure 1E).
Quantification and statistical analysis
All statistical analyses were performed by using GraphPad Prism 8 software. Graph data are presented as mean ± SD. Statistical details of the analyses can be found in the figure legends.
Acknowledgments
We thank Y. Kunihiro, T. Omoto, and S. Kobori at Osaka University and M. Hoshi and A. Fukui at Tokyo University, and S. Saji, S. Yamamoto, M. Omatsu, and N, Godai at RIKEN SPring-8 center for their technical assistance. This project was supported in part by JSPS KAKENHI Grant Numbers 18H03974 (T.M.) and 19K16025 (K.Y.), by the COVID-19 Emergency Action Fund from The University of Tokyo, and by the Platform Project for Supporting Drug Discovery and Life Science Research (JP20am0101070). This research was also partially supported by Platform Project for Supporting Drug Discovery and Life Science Research (Basis for Supporting Innovative Drug Discovery and Life Science Research [BINDS]) from AMED under Grant Number JP20am0101070 (support number 1251 and 2463), by the Research Program on Emerging and Re-emerging Infectious Diseases from AMED (19fk0108151 and JP19fk0108113), and by the National Institutes of Allergy and Infectious Diseases funded Center for Research on Influenza Pathogenesis (CRIP; HHSN272201400008C). We thank Sandra Cheesman, PhD, from Edanz (https://jp.edanz.com/ac), for editing a draft of this manuscript.
Author contributions
K.Y. designed and performed most of the experiments and analyzed the data with assistance from S.S. and Y.Y. K.T. and M.Y. prepared the CRISPR-Cas proteins and performed the experiments. S.Y., H.Y., and. Y.K. prepared the clinical samples for CRISPR diagnostic testing. T.M. conceived and supervised the study, prepared the figures, and wrote the manuscript with editorial contributions from all the authors.
Declaration of interests
S.S. is an employee of C4U. K.Y. and T.M. are cofounders of C4U and K.Y., K.T., and T.M. are a member of its scientific advisory board. K.Y., K.T., S.S., and T.M. are listed as inventors for a patent of CONAN-based detection assays.
Published: February 18, 2022
Footnotes
Supplemental information can be found online at https://doi.org/10.1016/j.isci.2022.103830.
Supplemental information
Data and code availability
This study did not generate any unique datasets or code. Any additional information required to reanalyze the data reported in this work paper is available from the Lead Contact upon request.
References
- Abbott T.R., Dhamdhere G., Liu Y., Lin X., Goudy L., Zeng L., Chemparathy A., Chmura S., Heaton N.S., Debs R., et al. Development of CRISPR as an antiviral strategy to combat SARS-CoV-2 and influenza. Cell. 2020;181:865–876.e12. doi: 10.1016/j.cell.2020.04.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ackerman C.M., Myhrvold C., Thakku S.G., Freije C.A., Metsky H.C., Yang D.K., Ye S.H., Boehm C.K., Kosoko-Thoroddsen T.F., Kehe J., et al. Massively multiplexed nucleic acid detection using Cas13. Nature. 2020;582:277–282. doi: 10.1038/s41586-020-2279-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beloglazova N., Petit P., Flick R., Brown G., Savchenko A., Yakunin A.F. Structure and activity of the Cas3 HD nuclease MJ0384, an effector enzyme of the CRISPR interference. EMBO J. 2011;30:4616–4627. doi: 10.1038/emboj.2011.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broughton J.P., Deng X., Yu G., Fasching C.L., Servellita V., Singh J., Miao X., Streithorst J.A., Granados A., Sotomayor-Gonzalez A., et al. CRISPR-Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 2020;38:870–874. doi: 10.1038/s41587-020-0513-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bustin S.A., Mueller R. Real-time reverse transcription PCR (qRT-PCR) and its potential use in clinical diagnosis. Clin. Sci. 2005;109:365–379. doi: 10.1042/CS20050086. [DOI] [PubMed] [Google Scholar]
- Chen J.S., Ma E., Harrington L.B., Da Costa M., Tian X., Palefsky J.M., Doudna J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science. 2018;360:436–439. doi: 10.1126/science.aar6245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chowdhury S., Carter J., Rollins M.F., Golden S.M., Jackson R.N., Hoffmann C., Nosaka L., Bondy-Denomy J., Maxwell K.L., Davidson A.R., et al. Structure reveals mechanisms of viral suppressors that intercept a CRISPR RNA-guided surveillance complex. Cell. 2017;169:47–57.e11. doi: 10.1016/j.cell.2017.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corman V.M., Landt O., Kaiser M., Molenkamp R., Meijer A., Chu D.K., Bleicker T., Brunink S., Schneider J., Schmidt M.L., et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill. 2020;25:2000045. doi: 10.2807/1560-7917.ES.2020.25.3.2000045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding X., Yin K., Li Z., Liu C. All-in-One dual CRISPR-Cas12a (AIOD-CRISPR) assay: a case for rapid, ultrasensitive and visual detection of novel coronavirus SARS-CoV-2 and HIV virus. bioRxiv. 2020 doi: 10.1101/2020.03.19.998724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dolan A.E., Hou Z., Xiao Y., Gramelspacher M.J., Heo J., Howden S.E., Freddolino P.L., Ke A., Zhang Y. Introducing a spectrum of long-range genomic deletions in human embryonic stem cells using type I CRISPR-Cas. Mol. Cell. 2019;74:936–950 e935. doi: 10.1016/j.molcel.2019.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forster P., Forster L., Renfrew C., Forster M. Phylogenetic network analysis of SARS-CoV-2 genomes. Proc. Natl. Acad. Sci. U S A. 2020;117:9241–9243. doi: 10.1073/pnas.2004999117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francois P., Tangomo M., Hibbs J., Bonetti E.J., Boehme C.C., Notomi T., Perkins M.D., Schrenzel J. Robustness of a loop-mediated isothermal amplification reaction for diagnostic applications. FEMS Immunol. Med. Microbiol. 2011;62:41–48. doi: 10.1111/j.1574-695X.2011.00785.x. [DOI] [PubMed] [Google Scholar]
- Fu B.X.H., Smith J.D., Fuchs R.T., Mabuchi M., Curcuru J., Robb G.B., Fire A.Z. Target-dependent nickase activities of the CRISPR-Cas nucleases Cpf1 and Cas9. Nat. Microbiol. 2019;4:888–897. doi: 10.1038/s41564-019-0382-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gootenberg J.S., Abudayyeh O.O., Kellner M.J., Joung J., Collins J.J., Zhang F. Multiplexed and portable nucleic acid detection platform with Cas13, Cas12a, and Csm6. Science. 2018;360:439–444. doi: 10.1126/science.aaq0179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gootenberg J.S., Abudayyeh O.O., Lee J.W., Essletzbichler P., Dy A.J., Joung J., Verdine V., Donghia N., Daringer N.M., Freije C.A., et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science. 2017;356:438–442. doi: 10.1126/science.aam9321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green D.A., StGeorge K. Rapid antigen tests for influenza: rationale and significance of the FDA reclassification. J. Clin. Microbiol. 2018;56 doi: 10.1128/JCM.00711-18. e00711–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo T.W., Bartesaghi A., Yang H., Falconieri V., Rao P., Merk A., Eng E.T., Raczkowski A.M., Fox T., Earl L.A., et al. Cryo-EM structures reveal mechanism and inhibition of DNA targeting by a CRISPR-Cas surveillance complex. Cell. 2017;171:414–426.e412. doi: 10.1016/j.cell.2017.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatakeyama S., Sugaya N., Ito M., Yamazaki M., Ichikawa M., Kimura K., Kiso M., Shimizu H., Kawakami C., Koike K., et al. Emergence of influenza B viruses with reduced sensitivity to neuraminidase inhibitors. JAMA. 2007;297:1435–1442. doi: 10.1001/jama.297.13.1435. [DOI] [PubMed] [Google Scholar]
- Hayes R.P., Xiao Y., Ding F., van Erp P.B., Rajashankar K., Bailey S., Wiedenheft B., Ke A. Structural basis for promiscuous PAM recognition in type I-E Cascade from E. coli. Nature. 2016;530:499–503. doi: 10.1038/nature16995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hidalgo-Cantabrana C., Barrangou R. Characterization and applications of Type I CRISPR-Cas systems. Biochem. Soc. Trans. 2020;48:15–23. doi: 10.1042/BST20190119. [DOI] [PubMed] [Google Scholar]
- Hitchman R.B., Possee R.D., King L.A. Baculovirus expression systems for recombinant protein production in insect cells. Recent Pat Biotechnol. 2009;3:46–54. doi: 10.2174/187220809787172669. [DOI] [PubMed] [Google Scholar]
- Hochstrasser M.L., Taylor D.W., Bhat P., Guegler C.K., Sternberg S.H., Nogales E., Doudna J.A. CasA mediates Cas3-catalyzed target degradation during CRISPR RNA-guided interference. Proc. Natl. Acad. Sci. U S A. 2014;111:6618–6623. doi: 10.1073/pnas.1405079111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imai M., Yamashita M., Sakai-Tagawa Y., Iwatsuki-Horimoto K., Kiso M., Murakami J., Yasuhara A., Takada K., Ito M., Nakajima N., et al. Influenza A variants with reduced susceptibility to baloxavir isolated from Japanese patients are fit and transmit through respiratory droplets. Nat. Microbiol. 2020;5:27–33. doi: 10.1038/s41564-019-0609-0. [DOI] [PubMed] [Google Scholar]
- Jackson R.N., Golden S.M., van Erp P.B., Carter J., Westra E.R., Brouns S.J., van der Oost J., Terwilliger T.C., Read R.J., Wiedenheft B. Structural biology. Crystal structure of the CRISPR RNA-guided surveillance complex from Escherichia coli. Science. 2014;345:1473–1479. doi: 10.1126/science.1256328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jore M.M., Lundgren M., van Duijn E., Bultema J.B., Westra E.R., Waghmare S.P., Wiedenheft B., Pul U., Wurm R., Wagner R., et al. Structural basis for CRISPR RNA-guided DNA recognition by Cascade. Nat. Struct. Mol. Biol. 2011;18:529–536. doi: 10.1038/nsmb.2019. [DOI] [PubMed] [Google Scholar]
- Kellner M.J., Koob J.G., Gootenberg J.S., Abudayyeh O.O., Zhang F. SHERLOCK: nucleic acid detection with CRISPR nucleases. Nat. Protoc. 2019;14:2986–3012. doi: 10.1038/s41596-019-0210-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimball A., Hatfield K.M., Arons M., James A., Taylor J., Spicer K., Bardossy A.C., Oakley L.P., Tanwar S., Chisty Z., et al. Asymptomatic and presymptomatic SARS-CoV-2 infections in residents of a long-term care skilled nursing facility - King County, Washington, March 2020. MMWR Morb Mortal Wkly Rep. 2020;69:377–381. doi: 10.15585/mmwr.mm6913e1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korber B., Fischer W., Gnanakaran S., Yoon H., Theiler J., Abfalterer W., Foley B., Giorgi E., Bhattacharya T., Parker M., et al. Spike mutation pipeline reveals the emergence of a more transmissible form of SARS-CoV-2. bioRxiv. 2020 doi: 10.1101/2020.04.29.069054. [DOI] [Google Scholar]
- Kumar S., Henrickson K.J. Update on influenza diagnostics: lessons from the novel H1N1 influenza A pandemic. Clin. Microbiol. Rev. 2012;25:344–361. doi: 10.1128/CMR.05016-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loeff L., Brouns S.J.J., Joo C. Repetitive DNA reeling by the Cascade-Cas3 complex in nucleotide unwinding steps. Mol. Cell. 2018;70:385–394 e383. doi: 10.1016/j.molcel.2018.03.031. [DOI] [PubMed] [Google Scholar]
- Lucia C., Federico P.-B., Alejandra G.C. An ultrasensitive, rapid, and portable coronavirus SARS-CoV-2 sequence detection method based on CRISPR-Cas12. bioRxiv. 2020 doi: 10.1101/2020.02.29.971127. [DOI] [Google Scholar]
- Matsuyama S., Nao N., Shirato K., Kawase M., Saito S., Takayama I., Nagata N., Sekizuka T., Katoh H., Kato F., et al. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. U S A. 2020;117:7001–7003. doi: 10.1073/pnas.2002589117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKimm-Breschkin J.L. Influenza neuraminidase inhibitors: antiviral action and mechanisms of resistance. Influenza Other Respir. Viruses. 2013;7:25–36. doi: 10.1111/irv.12047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metsky H.C., Freije C.A., Kosoko-Thoroddsen T.-S.F., Sabeti P.C., Myhrvold C. CRISPR-based surveillance for COVID-19 using genomically-comprehensive machine learning design. bioRxiv. 2020 doi: 10.1101/2020.02.26.967026. [DOI] [Google Scholar]
- Morisaka H., Yoshimi K., Okuzaki Y., Gee P., Kunihiro Y., Sonpho E., Xu H., Sasakawa N., Naito Y., Nakada S., et al. CRISPR-Cas3 induces broad and unidirectional genome editing in human cells. Nat. Commun. 2019;10:5302. doi: 10.1038/s41467-019-13226-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulepati S., Bailey S. Structural and biochemical analysis of nuclease domain of clustered regularly interspaced short palindromic repeat (CRISPR)-associated protein 3 (Cas3) J. Biol. Chem. 2011;286:31896–31903. doi: 10.1074/jbc.M111.270017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulepati S., Bailey S. In vitro reconstitution of an Escherichia coli RNA-guided immune system reveals unidirectional, ATP-dependent degradation of DNA target. J. Biol. Chem. 2013;288:22184–22192. doi: 10.1074/jbc.M113.472233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulepati S., Heroux A., Bailey S. Structural biology. Crystal structure of a CRISPR RNA-guided surveillance complex bound to a ssDNA target. Science. 2014;345:1479–1484. doi: 10.1126/science.1256996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murugan K., Seetharam A.S., Severin A.J., Sashital D.G. CRISPR-Cas12a has widespread off-target and dsDNA-nicking effects. J. Biol. Chem. 2020;295:5538–5553. doi: 10.1074/jbc.RA120.012933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Notomi T., Okayama H., Masubuchi H., Yonekawa T., Watanabe K., Amino N., Hase T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000;28:E63. doi: 10.1093/nar/28.12.e63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piepenburg O., Williams C.H., Stemple D.L., Armes N.A. DNA detection using recombination proteins. PLoS Biol. 2006;4:e204. doi: 10.1371/journal.pbio.0040204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ravina R., Dalal A., Mohan H., Prasad M., Pundir C.S. Detection methods for influenza A H1N1 virus with special reference to biosensors: a review. Biosci. Rep. 2020;40 doi: 10.1042/BSR20193852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redding S., Sternberg S.H., Marshall M., Gibb B., Bhat P., Guegler C.K., Wiedenheft B., Doudna J.A., Greene E.C. Surveillance and processing of foreign DNA by the Escherichia coli CRISPR-Cas system. Cell. 2015;163:854–865. doi: 10.1016/j.cell.2015.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rutkauskas M., Sinkunas T., Songailiene I., Tikhomirova M.S., Siksnys V., Seidel R. Directional R-loop formation by the CRISPR-Cas surveillance complex Cascade provides efficient off-target site rejection. Cell Rep. 2015;10:1534–1543. doi: 10.1016/j.celrep.2015.01.067. [DOI] [PubMed] [Google Scholar]
- Santiago-Frangos A., Hall L.N., Nemudraia A., Nemudryi A., Krishna P., Wiegand T., Wilkinson R.A., Snyder D.T., Hedges J.F., Cicha C., et al. Intrinsic signal amplification by type III CRISPR-Cas systems provides a sequence-specific SARS-CoV-2 diagnostic. Cell Rep. Med. 2021;2:100319. doi: 10.1016/j.xcrm.2021.100319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sashital D.G., Wiedenheft B., Doudna J.A. Mechanism of foreign DNA selection in a bacterial adaptive immune system. Mol. Cell. 2012;46:606–615. doi: 10.1016/j.molcel.2012.03.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato M., Takashita E., Katayose M., Nemoto K., Sakai N., Hashimoto K., Hosoya M. Detection of variants with reduced baloxavir marboxil susceptibility after treatment of children with influenza A during the 2018/2019 influenza season. J. Infect. Dis. 2020;222:121–125. doi: 10.1093/infdis/jiaa061. [DOI] [PubMed] [Google Scholar]
- Schneider C.A., Rasband W.S., Eliceiri K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 2012;9:671–675. doi: 10.1038/nmeth.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinkunas T., Gasiunas G., Fremaux C., Barrangou R., Horvath P., Siksnys V. Cas3 is a single-stranded DNA nuclease and ATP-dependent helicase in the CRISPR/Cas immune system. EMBO J. 2011;30:1335–1342. doi: 10.1038/emboj.2011.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teng F., Guo L., Cui T., Wang X.G., Xu K., Gao Q., Zhou Q., Li W. CDetection: CRISPR-Cas12b-based DNA detection with sub-attomolar sensitivity and single-base specificity. Genome Biol. 2019;20:132. doi: 10.1186/s13059-019-1742-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theel E.S., Slev P., Wheeler S., Couturier M.R., Wong S.J., Kadkhoda K. The role of antibody testing for SARS-CoV-2: is there one? J. Clin. Microbiol. 2020;58 doi: 10.1128/JCM.00797-20. e00797–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Udugama B., Kadhiresan P., Kozlowski H.N., Malekjahani A., Osborne M., Li V.Y.C., Chen H., Mubareka S., Gubbay J.B., Chan W.C.W. Diagnosing COVID-19: the disease and tools for detection. ACS Nano. 2020;14:3822–3835. doi: 10.1021/acsnano.0c02624. [DOI] [PubMed] [Google Scholar]
- van Erp P.B.G., Patterson A., Kant R., Berry L., Golden S.M., Forsman B.L., Carter J., Jackson R.N., Bothner B., Wiedenheft B. Conformational dynamics of DNA binding and Cas3 recruitment by the CRISPR RNA-guided Cascade complex. ACS Chem. Biol. 2018;13:481–490. doi: 10.1021/acschembio.7b00649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westra E.R., van Erp P.B., Kunne T., Wong S.P., Staals R.H., Seegers C.L., Bollen S., Jore M.M., Semenova E., Severinov K., et al. CRISPR immunity relies on the consecutive binding and degradation of negatively supercoiled invader DNA by Cascade and Cas3. Mol. Cell. 2012;46:595–605. doi: 10.1016/j.molcel.2012.03.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiedenheft B., Lander G.C., Zhou K., Jore M.M., Brouns S.J.J., van der Oost J., Doudna J.A., Nogales E. Structures of the RNA-guided surveillance complex from a bacterial immune system. Nature. 2011;477:486–489. doi: 10.1038/nature10402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao Y., Luo M., Dolan A.E., Liao M., Ke A. Structure basis for RNA-guided DNA degradation by Cascade and Cas3. Science. 2018;361:eaat0839. doi: 10.1126/science.aat0839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao Y., Luo M., Hayes R.P., Kim J., Ng S., Ding F., Liao M., Ke A. Structure basis for directional R-loop formation and substrate handover mechanisms in type I CRISPR-Cas system. Cell. 2017;170:48–60 e11. doi: 10.1016/j.cell.2017.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue C., Zhu Y., Zhang X., Shin Y.K., Sashital D.G. Real-time observation of target search by the CRISPR surveillance complex cascade. Cell Rep. 2017;21:3717–3727. doi: 10.1016/j.celrep.2017.11.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshimi K., Takeshita K., Kodera N., Shibumura S., Yamauchi Y., Omatsu M., Kunihiro Y., Yamamoto M., Mashimo T. Dynamic mechanisms of CRISPR interference by Escherichia coli CRISPR-Cas3. bioRxiv. 2021 doi: 10.1101/2021.07.18.452824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng Y., Li J., Wang B., Han J., Hao Y., Wang S., Ma X., Yang S., Ma L., Yi L., Peng W. Endogenous type I CRISPR-Cas: from foreign DNA defense to prokaryotic engineering. Front. Bioeng. Biotechnol. 2020;8:62. doi: 10.3389/fbioe.2020.00062. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
This study did not generate any unique datasets or code. Any additional information required to reanalyze the data reported in this work paper is available from the Lead Contact upon request.