

Abstract
The selective trafficking of proteins and RNAs through the nuclear envelope regulates nuclear-cytoplasmic segregation of macromolecules and is mediated by nucleopore complexes (NPCs), which consist of about 400 nucleoporins (Nups) of about 30 types. Extensive studies of nucleoporin function in yeast and vertebrates showed that Nups function in nucleocytoplasmic trafficking and other processes. However, limited studies of plant Nups have identified only a few mutations, which cause pleiotropic phenotypes including reduced growth and early flowering. Here, we describe loss-of-function alleles of Arabidopsis TRANSCURVATA1 (TCU1); these mutations cause increased hypocotyl and petiole length, reticulate and asymmetrically epinastic leaf laminae of reduced size, and early flowering. TCU1 is transcribed in all of the organs and tissues examined, and encodes the putative ortholog of yeast and vertebrate Nup58, a nucleoporin of the Nup62 subcomplex. Nup58 forms the central channel of the NPC and acts directly in translocation of proteins through the nuclear envelope in yeast and vertebrates. Yeast two-hybrid (Y2H) assays identified physical interactions between TCU1/NUP58 and 34 proteins, including nucleoporins, SCF (Skp1/Cul1/F-box) ubiquitin ligase complex components and other nucleoplasm proteins. Genetic interactions were also found between TCU1 and genes encoding nucleoporins, soluble nuclear transport receptors and components of the ubiquitin-proteasome and auxin signaling pathways. These genetic and physical interactions indicate that TCU1/NUP58 is a member of the Nup62 subcomplex of the Arabidopsis NPC. Our findings also suggest regulatory roles for TCU1/NUP58 beyond its function in nucleocytoplasmic trafficking, a hypothesis that is supported by the Y2H and genetic interactions that we observed.
Introduction
In eukaryotes, the nuclear envelope spatially separates certain key cellular processes, segregating them to the nucleus or the cytoplasm. This segregation depends on controlled nucleocytoplasmic traffic mediated by nucleopore complexes (NPCs), which allow regulated transport of macromolecules and passive diffusion of ions and small molecules [1]. The NPCs of Saccharomyces cerevisiae and vertebrates include multiple copies of about 30 different nucleoporins (Nups). Nups have been classified into three categories: (a) transmembrane Nups, which bind the NPC to the nuclear envelope, (b) FG-Nups, which contain phenylalanine-glycine (FG) repeats and (c) structural Nups, connecting transmembrane Nups to FG-Nups [2], [3]. FG-Nups are assumed to play a pivotal role in nuclear transport, acting as a selective permeability barrier and mediating the translocation of soluble nuclear transport receptors. In addition to the NPC, two types of soluble nuclear transport receptors also participate in nucleocytoplasmic traffic: nuclear export factors, which are involved in the exit of mRNA from the nucleus, and β-karyopherin family proteins, including importins and exportins, which transport proteins containing nuclear localization signals (NLSs) or nuclear export signals (NESs), respectively [4]. Nuclear import or export of proteins largely depends on their interaction with soluble nuclear transport factors, which recognize and bind the NLS or NES to their cargoes. Upon direct or indirect binding of a protein to its soluble nuclear receptor, the receptor and cargo are translocated together across the central channel of the nucleopore [5].
Little experimental evidence is available on nucleoporin function in plants. Several Arabidopsis nucleoporins have been identified based on similarity to their putative orthologs in yeast and vertebrates. The Arabidopsis orthologs of Nup155, Nup98, TPR (translocated promoter region) and GP210 (nuclear pore glycoprotein of 210 kDa) nucleoporins were identified in this way [6]. Other Arabidopsis nucleoporins, including WIP1 (WPP-domain Interacting Protein1), WIP2, WIP3 and NUA (Nuclear Pore Anchor, the Arabidopsis ortholog of TPR) were first identified in silico in a search for proteins with a coiled-coil coverage of at least 50% and either a NLS or at least one predicted transmembrane domain [7]; the WIP proteins were then found to colocalize at the nuclear envelope [8].
Mutant screens have also led to the identification of nucleoporins in Arabidopsis. For several Arabidopsis genes encoding nucleoporins, loss-of-function mutations cause pleiotropic phenotypes that include reduced growth and early flowering. In vertebrates, the Nup107–160 nucleopore subcomplex comprises Nup96, Nup107, Nup133 and Nup160. The Arabidopsis genes encoding NUP96 and NUP160 have been identified in several mutant screens performed with diverse objectives. For example, the mos3 (modifier of snc1, 3) and mos7 mutants were isolated in a screen for suppressors of snc1 (suppressor of npr1-1, constitutive1), a mutation that causes constitutive activation of disease resistance responses [9]. MOS3 encodes NUP96 and mos3-1 mutants exhibit small rosettes and early flowering. MOS7 encodes NUP88, which is required for innate immunity and nuclear accumulation of defense regulators [10]. The Arabidopsis sar1 (suppressor of auxin resistance1) and sar3 mutants were identified in a search for suppressors of the auxin resistance phenotype of axr1 (auxin resistance1). Additional sar1 alleles were isolated in a search for mutations impairing the cold-induced transcription of the CBF3-LUC (C-repeat/DRE Binding Factor 1-Luciferase) reporter gene [11]. SAR1 encodes NUP160, and sar3 mutations are allelic to mos3. Single sar1 and sar3 and double sar1 sar3 mutants display altered leaf morphology, early flowering, reduced rosette size and cell division rate in the primary root, and nuclear poly(A)+ RNA retention [12], [13].
In yeast, the TREX-2 complex (Transcription-coupled export 2) is anchored to the inner region of the NPC through Nup1, and is essential for mRNA export [14]-[18]. The TREX-2 complex includes the Thp1, Sac3, Sus1 and Cdc31 proteins. A yeast-two-hybrid (Y2H) screen using THP1 (Tho2/Hpr1 Phenotype) as bait identified a putative Arabidopsis ortholog of yeast Nup1, as well as other nucleoporins [19]. Mass-spectrometry analysis of proteins that coprecipitate with the RAE1 (RNA export factor 1) nucleoporin identified 30 components of the Arabidopsis NPC, of which 8 were previously described [20].
The central channel of the mammalian and yeast nucleopores is formed by the Nup54, Nup58 and Nup62 FG-nucleoporins of the Nup62 subcomplex, which in mammals also includes Nup45, a splice variant of Nup58. The Nup62 subcomplex is directly involved in the traffic of macromolecules across the nuclear envelope [21]. Here, we report the positional cloning of the TRANSCURVATA1 (TCU1) gene of Arabidopsis, which encodes a Nup58 ortholog. The perinuclear localization of TCU1, the phenotypic effects of tcu1 mutations, and the phenotypes of their double mutant combinations with alleles of genes involved in nucleocytoplasmic trafficking support that TCU1 is a nucleoporin. A Y2H-based screen of Arabidopsis cDNA libraries allowed us to identify TCU1 interactors including NUP62 and other nuclear proteins, some of which are components of the ubiquitin-proteasome and auxin signaling pathways.
Materials and Methods
Plant Materials and Growth Conditions
Seeds of the Arabidopsis thaliana L. Heynh. wild-type accessions Ler and Col-0 were obtained from the Nottingham Arabidopsis Stock Centre (NASC). The tcu1-1 mutant was isolated in the Ler background after EMS mutagenesis [22]. Seeds of T-DNA insertion lines were provided by the NASC or ABRC (Table S1) and are described at SIGnAL (http://signal.salk.edu) [23]. Seed sterilization and sowing, plant culture and crosses were performed as previously described [22], [24]. Briefly, seeds were sown on plates containing MS agar medium (half-strength Murashige and Skoog salts, 0.7% plant agar [Duchefa], pH 5.7, and 1% sucrose) and stratified (4°C in the dark) for 48 h and then transferred to either Conviron TC16 or TC30 growth chambers set to standard conditions of continuous light at approximately 75 µmol·m−2·s−1, 20°C, 60–70% relative humidity. When required, plants were transferred into pots containing a 2∶2∶1 mixture of perlite:vermiculite:sphagnum moss and grown in walk-in growth chambers set to the same standard conditions.
Plant Gross Morphology, Histology and Histochemical Assays
Leaf clearing and fixation, embedding, microscopy and morphometry were performed as previously described [25]–[28]. Venation, leaf epidermal and mesophyll cell diagrams were obtained from micrographs by hand drawing on the screen of a Wacom Cintiq 18SX Interactive pen display (http://www.wacom.com/) and using the Adobe Photoshop CS3 (http://www.adobe.com) software. Morphometric analyses of the diagrams (n≥10) were performed with ImageJ 1.36b [29] (http://rsb.info.nih.gov/ij/index.html/), Scion Image 4.0.3.2 and NIS-Elements AR 2.30 (Nikon Imaging; http://www.nis-elements.com/). GUS assays were performed as described in [30]. Unless otherwise indicated, all values in this paper are reported as means ± standard deviation from at least 10 (for morphometry) or 20 (all other experiments) plants.
Positional Cloning and Molecular Characterization of TCU1 and its Mutant Alleles
Low-resolution mapping of the tcu1-1 mutation was performed as previously described [31], [32]. For the fine mapping of the TCU1 gene, SSLP, SNP and In/Del markers were developed based on the polymorphisms between Ler and Col-0 described in the Monsanto Arabidopsis Polymorphism Collection database (http://www.arabidopsis.org). Synthetic oligonucleotides were purchased from Sigma-Aldrich UK (Table S2). Genomic DNA was extracted, PCR amplified and sequenced as described previously [33]. For the sequencing of tcu1 alleles, PCR amplification products spanning the At4g37130 transcription unit were obtained using wild-type and mutant genomic DNA as templates. To confirm the presence and position of T-DNA inserts, DNA was extracted and PCR amplified. Sequencing reactions, RNA extractions and qRT-PCR amplifications were performed as described in Barrero et al. [34], using the primers shown in Table S3. For qRT-PCR, each reaction was made using three biological replicates, each with three technical replicates; the expression levels were normalized to the CT values obtained for the housekeeping gene OTC [35].
DNA Constructs
To complement the tcu1-1 mutation, a 3.2-kb Ler genomic fragment containing the TCU1 coding region and its promoter was PCR amplified using the oligonucleotides indicated in Table S3. The amplification product was cloned into the EcoRI and XbaI sites of pGreen0179 (http://www.pgreen.ac.uk/JIT/pG0179.htm). The resulting TCU1pro:TCU1 construct was mobilized into Agrobacterium tumefaciens C58C1-pSOUP cells and then transferred into Col-0, Ler and tcu1-1 plants.
Gateway (Invitrogen) entry and destination vectors were used to obtain all the remaining constructs used in this work using the oligonucleotide primers indicated in Table S3. We constructed the TCU1pro:GUS transgene to visualize the expression pattern of the TCU1 gene, TCU1pro:TCU1:GFP to visualize the subcellular localization of the TCU1 protein, and 2x35S:TCU1 and 2x35S:tcu1-1 to overexpress the TCU1 and tcu1-1 alleles. We PCR amplified genomic fragments containing the TCU1 promoter (443 bp from Ler) for TCU1pro:GUS; the TCU1 promoter and coding region, lacking its stop codon (2,7 kb from Ler) for TCU1pro:TCU1:GFP; and the whole TCU1 coding region (2,3 kb from either Ler or tcu1-1) for 2x35S:TCU1 and 2x35S:tcu1-1, respectively. The amplification products were cloned into the pGEM-T Easy221 vector, sequence verified, transferred into the pMDC164, pMDC111 and pMDC32 destination vectors, respectively, and sequence verified again. The resulting constructs were mobilized into Agrobacterium tumefaciens LBA4404 cells, and then transferred into Col-0, Ler, tcu1-1 and tcu1-2 plants. Arabidopsis plant transformations were performed by the floral dip method [36], and the transgenic plants were selected on MS agar medium supplemented with 15 µg·ml−1 hygromicin.
Ubiquitin Immunodetection
Protein extraction and immunoblotting were performed as described in [37]. Antibody (antiUBQ11; Agrisera) serum [38] was used at a dilution of 1∶1,000 (v/v). The immunoreactive proteins were visualized using Pierce picosignal reagents, with the secondary rabbit antibody diluted 1∶5,000 (v/v), and by exposure to X-ray film (Amersham Hyperfilm) for 10 s to 1 h.
Results
The tcu1-1 Mutation Causes a Pleiotropic Phenotype
We performed a large-scale screen for EMS-induced mutants with altered leaf morphology in the Ler genetic background. Some mutants exhibited leaves folded towards the abaxial surface in an asymmetrical manner relative to the primary vein. We called these mutants tcu (transcurvata). The tcu mutations are recessive and fully penetrant, with only small variations in expressivity, and fall into three complementation groups (TCU1, TCU2 and TCU3) [22].
The cotyledons of the tcu1-1 mutant failed to expand completely; also, vegetative leaf laminae were slightly smaller than wild type (Figure 1A-E), and reticulated, with veins greener than the interveinal regions. These visible traits correlated with perturbations in internal leaf anatomy, as shown by the enlarged air spaces and fewer cells in the interveinal spongy mesophyll seen in transverse sections (Figure 1F-H). The leaf veins and their perivascular bundle sheath cells seemed unaffected. Differential interference contrast (DIC) microscopy of first- and third-node tcu1-1 and Ler leaves cleared with chloral hydrate revealed only small differences in cell number or cell size in the abaxial and adaxial epidermal layers, as well as in the subepidermal layer of palisade mesophyll cells (Figure S1). Similar to vegetative leaves, cauline leaves were folded downwards.
Figure 1. Morphological and cellular phenotypes of the tcu1-1 mutant.

(A) Lateral view of seedlings and (B, C) top view of rosettes of the indicated genotypes. (D, E) From left to right, dissected cotyledons and first to ninth (in Ler) or seventh (in tcu1-1) vegetative leaves. (F-H) Leaf transverse sections and (I, J) hypocotyl cells from plants of the indicated genotypes. pv: primary vein. Asterisks in H highlight enlarged intercellular air spaces. Plants were collected at (A) 12, (B, C, F–J) 21 and (D, E) 22 das. Scale bars: (A) 1 mm, (B, C) 2 mm, (D, E) 5 mm, (F) 200 µm and (G–J) 50 µm.
Hypocotyls of plants grown in our standard conditions were longer in tcu1-1 (2.09±0.26 mm) than in Ler (1.46±0.27 mm). Hypocotyl epidermal cell length was 194.85±32.79 µm in the mutant and 91.30±19.08 µm in the wild type (Figure 1I, J). The petiole and its adaxial epidermal cells were longer in the tcu1-1 mutant (4.42±0.74 mm and 444.94±168.14 µm, respectively) than in Ler (3.09±0.46 mm and 320.41±134.40 µm) in first-node leaves collected 13 das (days after stratification). Bolting was observed at 21.86±0.94 das in tcu1-1 and 27.89±1.36 das in Ler, and the mutant developed fewer vegetative leaves (7.36±0.81) than Ler (12.40±0.52).
No differences with the wild type were observed in root, inflorescence, flower and fruit morphology, root gravitropism and skotomorphogenesis. The venation patterns of tcu1-1 leaves had slightly increased total vein length and number of free-ending veins, but reduced number of branching points compared with Ler (Figure S2 and Table S4).
The TCU1 Gene Encodes the Arabidopsis NUP58 Nucleoporin
For the positional cloning of TCU1, we performed linkage analysis on an F2 mapping population of 740 plants derived from a tcu1-1× Col-0 cross (see Materials and Methods). This allowed us to delimit a 77-kb candidate region on chromosome 4, encompassing 19 annotated genes (At4g37070-At4g37260) (Figure 2A). Sequencing of these genes in the tcu1-1 mutant revealed a C→T transition predicted to create a premature stop codon at nucleotide position 184 (numbering from the predicted translation initiation codon) in At4g37130 (Figure 2B).
Figure 2. Positional cloning, structure and constructs of the TCU1 gene.

(A) Map-based cloning strategy. The molecular markers used for linkage analysis and the number of informative recombinants identified (in parentheses) are indicated. (B) Schematic representation of the TCU1 gene and tcu1 mutant alleles. Triangles represent T-DNA insertions. Exons are shown as boxes, and introns as lines between boxes. Open boxes indicate 5′ and 3′ untranslated regions. Translation start and stop codon positions are shown. Horizontal arrows represent oligonucleotides used as primers for qRT-PCR amplifications (not drawn to scale). (C) Span of the genomic DNA (red) or cDNA (black) segments amplified by PCR to obtain the constructs shown, which were used for the functional characterization of TCU1.
To confirm that loss of At4g37130 function causes the tcu1-1 mutant phenotype, we conducted several transgene-mediated complementation experiments. A wild-type (Ler) genomic segment encompassing the entire region between At4g37120 and At4g37140 in the TCU1pro:TCU1 transgene (Figure 2C) was transferred into tcu1-1 plants, and all the resulting transformants displayed wild-type morphology (Figure S3). Another transgene including the full coding region of At4g37130 expressed under the control of the 35S promoter (2x35Spro:TCU1; Figure 2C) also rescued the phenotype of the tcu1-1 mutant but had no phenotypic effects in the Ler background. A construct carrying the same genomic segment of the mutant allele (2x35Spro:tcu1-1) did not cause any phenotypic effects in the Ler or tcu1-1 backgrounds.
Four publicly available lines carrying T-DNA insertions within the At4g37130 gene were identified (Table S1). Their insertions were sequence-confirmed to be at the annotated nucleotide positions. Allelism tests further confirmed that they carry TCU1 alleles, which we called tcu1-2 to tcu1-5. All these insertional lines shared phenotypic traits with tcu1-1 to different degrees, and these shared phenotypes include early flowering (Figure 3) and increased petiole length. Leaf lamina folding and reduced size were similar in tcu1-1 and tcu1-4 mutants, but milder in the remaining lines.
Figure 3. Early flowering in the tcu1 mutants.
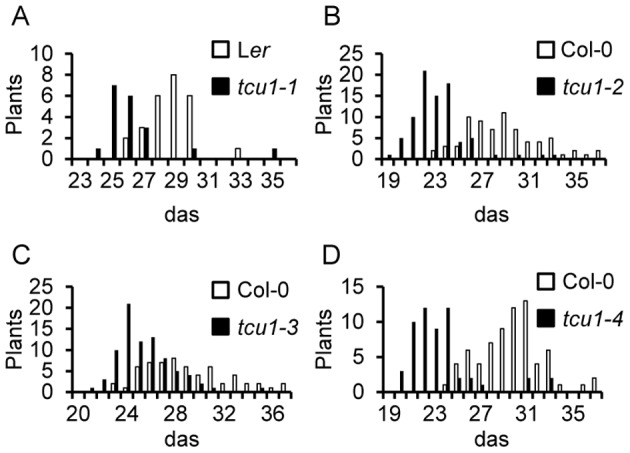
Bolting time of (A) tcu1-1 versus its wild type Ler, and (B–D) tcu1-2, tcu1-3 and tcu1-4 versus their wild type Col-0.
TCU1 is predicted to encode an FG-nucleoporin of 513 amino acids and a molecular mass of 56.5 kDa (http://www.arabidopsis.org/servlets/TairObject? id = 126619&type = locus), a putative member of a hydroxyproline-rich glycoprotein family, closely related to mammalian Nup58/45 (Figure S4). To further validate its identity as a nucleoporin, we determined the subcellular localization of TCU1 using a translational fusion to the Green Fluorescent Protein (GFP) marker, under the control of the native TCU1 promoter, TCU1pro:TCU1:GFP (Figure 2C). This transgene completely complemented the morphological phenotype of tcu1-1, indicating that the GFP fusion did not impair TCU1 function. As expected for a predicted nucleoporin, the GFP signal was detected only along the nuclear envelope in root and leaf mesophyll cells (Figure 4).
Figure 4. Subcellular localization of the TCU1:GFP protein and spatial pattern of expression of TCU1.
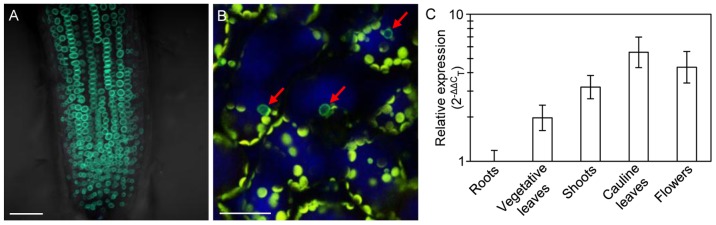
(A, B) Confocal images of cells from (A) the root elongation zone and (B) the subepidermal layer of leaf palisade mesophyll from Ler plants carrying the TCU1pro:GFP transgene. The green, perinuclear signal corresponds to GFP, and the yellow signal to chloroplast chlorophyll autofluorescence. Red arrows in B indicate nuclei. Scale bars: 20 µm. (C) qRT-PCR analysis of expression of TCU1. Total RNA isolated from the tissues indicated was used as a template. Error bars represent standard deviations.
TCU1 is Broadly Expressed
We examined in silico the expression profile of TCU1 using Genevestigator (http://www.genevestigator.ethz.ch) [39] and the Arabidopsis eFP Browser (http://bbc.botany.utoronto.ca/efp/cgi-bin/efpWeb.cgi) and found that TCU1 is expressed in all tissues and developmental stages in Arabidopsis (Figure S5). To confirm this expression pattern, we also performed real-time qRT-PCR amplifications using RNA from assorted tissues of Col-0 and Ler. The highest and lowest expression levels of TCU1 were detected in cauline leaves and roots, respectively (Figure 4C).
TCU1 expression was further analyzed by transferring a TCU1pro:GUS construct (Figure 2C) into Col-0, Ler, tcu1-1 and tcu1-2 plants, with similar results in all genetic backgrounds. GUS activity was detected in all the stages and tissues analyzed of 4, 7, 11, 15, 18 and 21 das transgenic plants (Figure S6), being most intense in leaf primordia and the vasculature. The highest expression levels were detected between 7 and 15 das.
TCU1 Physically Interacts with Nucleoporins and Other Nuclear Proteins, Including ASK1 and ASK2
To gain insight into the function of TCU1, the Y2H system was used to screen for TCU1 interactors. Two Arabidopsis cDNA libraries, totalizing 21 million prey clones, were screened at PanBioNet (http://www.panbionet.com) using the TCU1_93-513 bait, which encompasses amino acids 93 to 513 of TCU1. This screen identified ninety prey clones, representing 34 different genes, 17 of which encode known nuclear proteins (Table S5 and Figure S7).
Proteins particularly well-represented in the Y2H-based screen included GAI (GA INSENSITIVE, 5 clones), which is a member of the DELLA family of growth repressors [40]-[42] and key negative regulators of gibberellin signaling [43]. Other identified proteins included NUP62 (also named EMB2766, 4 clones); ACT7 (4 clones), a component of the actin pathway [44]; the AT1G21440 mutase (4 clones); and the AT3G20720 unknown protein (4 clones). In most other cases only one or two positive clones were identified for a given protein. However, the screen also identified 8 ASK1 and 15 ASK2 prey clones. SKP1 (S PHASE KINASE-ASSOCIATED PROTEIN1), also called ASK1 (ARABIDOPSIS SKP1-LIKE1), and ASK2 are components of the SKP1-cullin/CDC53-F-box protein (SCF) ubiquitin-protein ligase complex, which plays important roles in selecting substrates for proteolysis by facilitating the ligation of ubiquitin to specific proteins [45], [46]. This interaction of TCU1/NUP58 with two SCF components prompted us to determine the levels of ubiquitin conjugates in the tcu1 mutants. However, we found that the levels of high molecular mass polyubiquitinated proteins were not apparently altered in the tcu1 mutants compared to their wild types (Figure 5).
Figure 5. Ubiquitination in the tcu1 mutants.
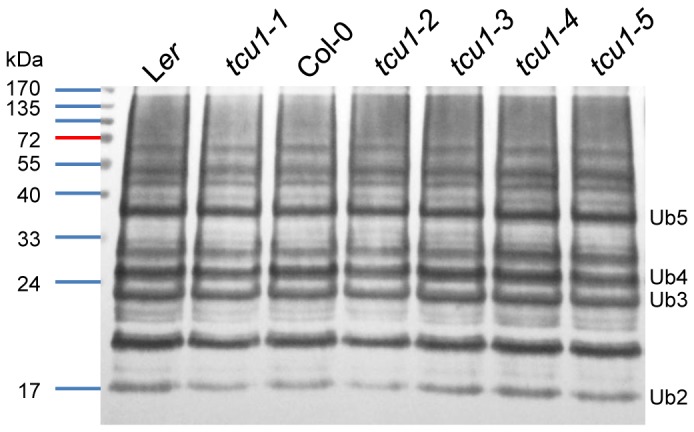
Twenty micrograms of protein was electrophoresed on a 5% stacking/10% separating polyacrylamide gel, electroblotted onto nitrocellulose membrane and probed with an anti-ubiquitin (antiUBQ11) antibody. The positions of molecular mass markers are shown on the left, and those of some poly-ubiquitin chains on the right.
TCU1 Genetically Interacts with Genes Required for Nucleocytoplasmic Trafficking and the Ubiquitin-proteasome Pathway
To examine the genetic interactions among genes encoding proteins involved in nucleocytoplasmic transport, we generated double mutants between tcu1-1 and tcu1-2 and loss-of-function mutations in genes encoding putative or known nucleoporins, soluble nuclear receptors and some of the interactors found in the Y2H screen (Table S5 and Figure S7).
The nup54-1 and nup62-1 mutations (Figure 6E, F), and nup54-2 and nup62-2 to a lesser extent, and also caused early flowering, increased petiole length and reduced lamina size in vegetative leaves. A synergistic phenotype was seen in all double mutant combinations of tcu1 alleles and nup54-1, nup54-2 or nup62-1 (Figure 6K-M), consisting of much smaller and darker rosettes than in their single mutant parental lines. A functional relationship between TCU1/NUP58, NUP54 and NUP62 was expected because they all belong to the Nup62 subcomplex. The tcu1-1 nup62-2 and tcu1-2 nup62-2 double mutants were pale and exhibited very long petioles (Figure 6N, O), phenotypes that were also considered to be synergistic.
Figure 6. Genetic interactions between tcu1 alleles and alleles of genes known or suspected to be involved in nucleocytoplasmic trafficking.

All plants are homozygous for the mutations indicated. Pictures were taken at (A, B, I-T) 21, (C, D) 22, (E, F) 25 and (G, H) 18 das. Scale bars: 2 mm.
We also observed synergistic phenotypes in the tcu1 sar1 and tcu1 sar3 double mutants (Figure 6P-R), an observation that indicates a functional relationship between TCU1 and genes encoding components of the Nup160 nucleopore subcomplex. Mutations in genes encoding soluble nuclear receptors such as hst-21 and imn interacted synergistically with tcu1-1 and tcu1-2. HST (HASTY) is the Arabidopsis ortholog of mammalian exportin5 and yeast MSN5, two importin β-like soluble transport receptors. The HST protein is thought to regulate the nucleocytoplasmic transport of microRNA molecules [47]. The phenotype of the hst-21 tcu1-1 double mutant differed from that of its single mutant parentals, with small rosettes, strongly reticulated leaves, short stems, flowers and fruits, and reduced fertility (Figure 6S). The strong hyponasty of hst-21 leaves was almost completely suppressed in the hst-21 tcu1-1 double mutant. The imn mutant carries a T-DNA insertion in the At5g53480 gene, which encodes a homolog of the human importin β1 [48]. The imn mutant exhibited generalized depigmentation, small rosettes and early flowering. The phenotype of the tcu1-1 imn double mutant was synergistic, with pale, small, narrow and pointed leaves, and long petioles (Figure 6T). The IMN protein was identified as an interactor of NUP136 using mass spectrometry [20], but has not been studied further. Taken together, these observations suggest a functional relationship between TCU1 and other NPC proteins.
We also crossed tcu1-1 to mutants carrying alleles of genes encoding components of the auxin signaling pathway. These included mutants in two Aux/IAA genes, the axr3-3 semidominant allele of AUXIN RESISTANT3 [49], [50], and the icu5 dominant allele of SHORT HYPOCOTYL2 (SHY2) [51]–[54]. We also crossed tcu1-1 to the recessive axr1-12 allele of AXR1, encoding a subunit of the RUB1 activating enzyme, which regulates the protein degradation activity of SCF complexes [55]. The tcu1-1 axr3-3 and tcu1-1 axr1-12 double homozygotes exhibited phenotypes intermediate between those of their tcu1-1 and axr3-3 single mutant siblings, and the tcu1-1 icu5 and icu5 siblings were indistinguishable (Figure 7). The observed partial mutual suppression of tcu1-1 and axr3-3, as well as that of tcu1-1 and axr1-12 suggests that TCU1 interacts with AXR3 and AXR1. The observation that icu5 is epistatic to tcu1-1 also suggests an interaction between SHY2 and TCU1.
Figure 7. Genetic interactions between tcu1-1 and alleles of genes encoding components of the auxin signaling pathway.

The genetic backgrounds of the plants shown are (A) En-2, (B) Col-0, (C) Col-1, (D) Ler, (E, H) Ler/En-2, (F, I) Ler/Col-0 and (G, J) Ler/Col-1. All plants are homozygous for the mutations indicated. Pictures were taken at 21 das. Scale bars: 1 mm.
The tcu1 Mutants are Hypersensitive to 2,4-D and Paclobutrazol and Defective in Light Perception
We further characterized the functions of TCU1 by examining the hormone responses in tcu1-1 mutants. To examine whether TCU1 plays a role in auxin responses, we tested whether the tcu1-1 mutant was hypersensitive to 2,4-D (2,4-dichlorophenoxyacetic acid). On medium containing 50 nM 2,4-D, root growth was reduced to 51.8% (compared to untreated roots) in the mutant and 95.05% in Ler. In addition, the tcu1-1 mutant was hypersensitive to the blockage of gibberellin synthesis caused by paclobutrazol: germination inhibition was stronger in tcu1-1 (22.7% germination) than in Ler (51.1% germination) in the presence of 1 µM paclobutrazol. We also studied the effect of different light intensities on the phenotype of tcu1-1 and tcu1-2 and found that it worsened under low light (40 µmol·m−2·s−1) but was almost completely suppressed by high light (130 µmol·m−2·s−1) (Figure S8).
Discussion
Most of our current knowledge on NPC structure and function derives from studies performed in yeast and animals. The Nup54 and Nup58 components of the Nup62 subcomplex were first identified from rat liver nuclei [56]. Later, Nup54, Nup58 and Nup62 were found to coprecipitate in a complex from rat and Xenopus protein nuclear extracts [57]. More recently, the crystal structures of the interacting domains of Rattus norvegicus Nup54, Nup58 and Nup62 were determined, indicating that as many as 224 copies of these proteins form a flexible transport channel with a ring diameter that can be modulated by circumferential sliding induced by interactions with transport receptors [58], [59].
The Nup62 subcomplex has received little attention in plants. Arabidopsis NUP54, NUP58 and NUP62 coimmunoprecipitate with RAE1-GFP, a GFP-tagged nucleoporin [20]. Although insertional alleles of the corresponding genes are available, no mutational analysis has been published, the only exception being the demonstration that overexpression-based cosuppression of the gene encoding NUP62 causes stunted growth and early flowering [60].
In this work, positional cloning, complementation and comparative analysis of five tcu1 mutant alleles confirmed that their phenotype is caused by loss of function of the At4g37130 gene, which encodes a likely ortholog of the Nup58 nucleoporin of yeast and vertebrates. All tcu1 alleles studied here are viable and recessive and their lesions most likely cause a partial or complete loss of function of TCU1. TCU1 is a single-copy gene, but does not seem to be essential in Arabidopsis, an idea supported by the viability of mutant plants carrying the tcu1-1 allele, which encodes a truncated protein with only 61 aa. Indeed, the gene encoding NUP62 is included in the database of Arabidopsis embryonic lethal mutants, but the genes encoding TCU1/NUP58 and NUP54 are not included, indicating that they also are not essential [61]. The TCU1pro:TCU1, 2x35Spro:TCU1 and 2x35Spro:tcu1-1 transgenes did not alter the phenotype in wild type backgrounds, indicating that an excess of TCU1/NUP58 function has no significant morphological effects and suggesting that the truncated protein produced by tcu1-1 is not antimorphic.
We have shown that TCU1 is expressed in all the tissues and organs studied, as would be expected for a putatively domestic function that is likely to be required in every cell. The spatial expression pattern of TCU1 is similar to those of other previously described genes encoding Arabidopsis nucleoporins, such as NUA [62] and SAR1 and SAR3 [13]. We have also shown that a functional TCU1:GFP fusion protein localizes to the nuclear envelope, in line with the results of Tamura et al. [20]. Perinuclear localization has also been demonstrated for other putative NPC components of Arabidopsis: SAR3/NUP96 [13], WIP1, WIP2a and WIP3 [8], NUA/TPR [7], and SEH1, SEC13, GP210, NUP54, NUP75, NUP88, NUP93a, NUP107, NUP1/NUP136, NUP160/SAR1, NUP205 and NUP214 [20].
The pleiotropic phenotype of tcu1 mutants suggests that TCU1 is required in several aspects of Arabidopsis development and physiology. The increased length of tcu1-1 petioles and hypocotyls results from increased cell elongation. The size reduction of the lamina of tcu1-1 appears to be due to a reduced number of cell divisions in both the epidermis and palisade mesophyll of vegetative leaves, whose cells show no appreciable changes in shape or size. A deficit in the number of leaf cells is particularly apparent in the spongy mesophyll, which exhibits large air spaces.
We also conducted a Y2H screen for TCU1/NUP58 interactors; this screen identified 34 genes, including 17 that encode known nuclear-localized proteins. An indicator of the quality of the screen is that one of the preys identified was NUP62. These results are consistent with our genetic interaction analyses, in which we combined alleles of TCU1 and genes encoding members of the Nup62 and Nup107–160 nucleopore subcomplexes, an exportin and an importin. The double mutant phenotypes were synergistic in all cases, indicating a functional relationship with TCU1/NUP58 and suggesting that TCU1 acts at the nucleopore and is involved in nucleocytoplasmic trafficking.
Some hormone signals were altered in the tcu1-1 mutant, which was hypersensitive to 2,4-D, a synthetic auxin, and paclobutrazol, an inhibitor of gibberellin synthesis. The hypersensitivity of the tcu1-1 mutant to paclobutrazol during germination suggests insufficient translocation to the nucleus of a positive regulator of the gibberellin pathway. AXR1 encodes an enzyme required for the activation of SCF ubiquitin ligase complexes [55], [63], [64]. Insensitivity to auxin in axr1 mutants is due to the nuclear accumulation of Aux/IAA proteins, which regulate auxin-responsive genes and are regulated by the SCFTIR1 complex (reviewed in [65]. The hypersensitivity to auxin in tcu1-1 mutants suggests that TCU1/NUP58 is required for the translocation of one or several SCFTIR1 complex components or Aux/IAA transcription factors to the nucleus. In line with this, Parry et al. [13] proposed that altered auxin homeostasis explains the phenotypes of sar1 and sar3 mutants. These authors hypothesized that altered NPC function impairs the nuclear import of an auxin signaling protein in sar1 and sar3 mutants. Although alterations in leaf vasculature are frequently found in mutants affected in auxin homeostasis, the venation pattern of the tcu1-1 mutant was relatively normal.
The mutant phenotype of tcu1 alleles is enhanced by reduced light intensities and suppressed by increased light. This observation suggests that TCU1/NUP58 is required for light perception. In addition, the phenotype of tcu1 mutants includes traits that suggest an altered photomorphogenic response: early flowering, leaf lamina size reduction, increased hypocotyl and petiole length, and acute angles of the petioles with the stem. These traits define the shade avoidance syndrome shown by plants growing under a canopy of other plants, which partially deprive them of sunlight (reviewed in [66], [67]). Solar radiation is rich in the red and blue components of the visible spectrum, which are partly absorbed by the plants exposed to direct sunlight, creating an area of shadow enriched in far-red wavelengths. Light rich in the far-red component is perceived by PHYB and other photoreceptors, activation of which triggers changes in the expression levels of many genes, which in turn gives rise to the morphological changes characterizing the shade avoidance syndrome [68]–[70]. Poor perception of visible light and, in particular, of its red component might explain the constitutive shade avoidance syndrome of tcu1 mutants, as well as its suppression by intense light.
Reduced Nup96 levels impair immune responses in mice, apparently due to reduced export of mRNAs encoding proteins regulating immunity [71]. In spite of the phylogenetic distance between mammals and plants, Nup96 and two other nucleoporins of the Nup107–160 subcomplex are also involved in immunity in Arabidopsis. A connection with defense has been demonstrated only for NUP96 (MOS3) [9], SEH1 and NUP160 [72], but not for the remaining five nucleoporins of the Nup107–160 complex, which are encoded by single-copy genes in the Arabidopsis genome and should not, in principle, be affected by redundancy. This observation supports several explanations, one of which is that some nucleoporins have specific functions, not necessarily related to their contribution to nucleocytoplasmic transport.
Comparisons among mutants affected in NPC function in Arabidopsis clearly indicate the existence of both common and specific phenotypic traits. Common traits suggest that the genes are involved in the same process. By contrast, the specific traits might be attributed to allele specificities when a single allele is available. However, two additional likely explanations are (1) that specific NPC components are particularly critical for the transport of specific molecules, and (2) that specific NPC components play roles beyond nuclear transport. The genetic and physical interactions described here are consistent with the hypothesis that TCU1/NUP58 is a member of the Nup62 subcomplex of the Arabidopsis NPC. Our findings also suggest regulatory roles for TCU1/NUP58 beyond its participation in nucleocytoplasmic trafficking. In this regard, it is worth noting experimental evidence obtained in yeast, Drosophila melanogaster and mammals, that indicates active role of nucleopore components in regulating gene expression. Some nucleoporins are suspected to act as insulators in human cells, separating active and inactive chromatin domains (reviewed in [73]. Studies in yeast and human cells indicate that nucleopore components are also involved in DNA repair and maintenance of genome integrity (reviewed in [74]). In addition, there are several nucleoporins of Drosophila melanogaster with a demonstrated function outside the nucleopore [75]: Nup88, Nup98 and Sec13 bind to chromatin and Nup98 and Sec13 act as transcription factors [76]; Nup153 is also involved in transcription [77], and Nup98, Nup62 Nup50 act together as transcription factors in embryonic cells [78].
A close relationship exists between nucleoporins and sumoylation, a process apparently not related to nucleo-cytoplasmic trafficking. Mutations in the NUA gene cause an increase of SUMO conjugates [7], similar to that caused by mutations in ESD4 (EARLY IN SHORT DAYS4), which encodes a nuclear protease that participates in sumoylation [79], [80]. NUA and ESD4 interact in Y2H assays [7]. Mutations in the gene encoding NUP160 also increase the levels of SUMO conjugates [57].
We have not studied the relationship between TCU1/NUP58 and sumoylation. However, the most represented preys among the positive clones identified in our Y2H screen were two components of the SCF complex: SKP1/ASK1 and ASK2. Since this observation suggested a connection between the TCU1/NUP58 nucleoporin and ubiquitination, we analyzed total protein extracts and found that ubiquitin conjugate levels are not different in the tcu1 mutants and their wild types. On the contrary, we found genetic interactions between tcu1 alleles and alleles of genes of the ubiquitin-proteasome pathway. In conclusion, the results of our Y2H analysis suggest that TCU1/NUP58 binds ASK2 and ASK1, and the results of our genetic analyses indicate a functional relationship between TCU1/NUP58 and auxin signaling, as well as with at least one component of the ubiquitin-proteasome pathway. We did not detect, however, any difference between the tcu1 mutants and their wild types in the levels of polyubiquitinated proteins. Further experiments will be required to ascertain the role of TCU1/NUP58 in ubiquitination.
Although we have not found any relationship between TCU1/NUP58 and proteasome components, it should be noted that two of the Arabidopsis homologs of components of the yeast TREX-2 complex, which is anchored to the NPC, interact with the Arabidopsis homolog of DSS1, which is an established proteasome component in yeast and animals. This observation suggests a link between the two complexes [19].
The tcu1 mutants may be instrumental in revealing the relationship between an apparently domestic cellular process, nucleocytoplasmic transport, and specific aspects of plant development such as light-regulated development, leaf organogenesis and flowering. Some of the observed physical and genetic interactions will require confirmation by other experimental approaches. The identified interactors indicate a possible relationship between TCU1/NUP58 and specific biological processes, including the ACT2 and ACT7 actins and development, KNAT3, HY2 and DET3 and light perception and signaling, and GAI and the flowering-promoting pathway mediated by gibberellins. Indeed, the response to gibberellin is known to be elicited through targeted degradation of DELLA proteins by the 26S proteasome via the SCFSLY complex [81].
Supporting Information
Leaf cellular phenotypes of tcu1-1 and Ler. (A–D) Adaxial epidermal cells shown as (A, B) interference contrast micrographs and (C, D) diagrams. (E, F) Diagrams of abaxial epidermal cells. Stomata are only partially drawn and appear as circles in C–F. (G, H) Diagrams of cells of the palisade mesophyll subepidermal layer. (I) Boxplot distribution of cell sizes in the tissues and genotypes shown. Boxes are delimited by the first (Q1, lower hinge) and third (Q3, upper hinge) quartiles. Whiskers represent Q1–1.5·IQ (lower) and Q3+1.5·IQ (upper), where IQ = Q3– Q1. ⋄: Mean. –: Median. ○: Extreme maximum outlier (> [Q3+3·IQ]). × : Maximum outlier. Leaves were collected at 21 das (days after stratification). Scale bars: 50 µm.
(PPTX)
Venation pattern in tcu1-1 leaves. Diagrams were drawn from first- and third-node leaves collected 21 das. The leaf margin is shown in orange. Some excisions at the margin were required to flatten tcu1-1 leaves before microscopy. Scale bar: 2 mm.
(PPTX)
Phenotypic complementation of tcu1-1 by the TCU1pro:TCU1 transgene. The plants shown were isolated on medium supplemented with 15 µg·ml−1 hygromicin among the T1 progeny of tcu1-1 plants transformed by infection with Agrobacterium tumefaciens C5851 cells carrying the pGreen0179 plasmid either without any insert (left) or with the TCU1pro:TCU1 insert (right). The picture was taken 26 das. Scale bar: 2 cm.
(PPTX)
Comparison of deduced amino acid sequences of TCU1 and the putative Nup58s of some higher plants. The TCU1/NUP58 protein of Arabidopsis thaliana (At; NP195430.2) is aligned with homologous gene products from Nicotiana tabacum (Nt; ACY30439.1), Vitis vinifera (Vv; XP_002282659), Populus balsamifera subsp. Trichocarpa (Pb; XP_002310761.1), Oryza sativa (Os; NP_001063345.1) and Zea mays (Zm; NP001132589). Amino acid residues identical or similar in all five sequences are shaded black or grey, respectively. The first of the 452 amino acids that are predicted to be missing in the tcu1-1 mutant is shaded red. The alignment was obtained using Clustal X 2.0 (Larkin et al., 2007) and shaded with Boxshade 3.21 (http://www.ch.embnet.org/software/BOX_form.html).
(PPTX)
Expression data output obtained from the Arabidopsis Electronic Fluorescent Pictograph (eFP) Browser for At4g37130 (TCU1) expression levels throughout all Arabidopsis developmental stages.
(PPTX)
Spatial expression analysis of TCU1. GUS staining of TCU1pro:GUS transgenic plants in (A) roots, (B) a cotyledon, (C–I) expanding leaves and whole rosettes. Plant material was collected at the time shown in each picture (in das). Scale bars: (A–C) 0.5 mm and (D–I) 1 mm.
(PPTX)
Confirmation of interactions identified in a Y2H screen with TCU1_93–513 as bait. Yeast PBN204 cells containing three reporters (URA3, lacZ, and ADE2) that are under the control of different GAL promoters were used. Yeast transformants of the TCU1_93–513 bait and 2 different Arabidopsis cDNA AD libraries were spread on SD-LWU (SD without leucine, tryptophan and uracil) selection medium, which supports growth of cells with bait and prey plasmids yielding proteins interacting each other. After selecting yeast colonies on uracil-deficient media, beta-galactosidase activity was monitored. Growth of the URA+ and lacZ+ colonies on adenosine-deficient media was also tested. This three independent reporter system reduces false positives. In order to confirm the interactions found, the prey parts of the plasmids of the positive clones were amplified by PCR and reintroduced into yeast, each with either the TCU1_93–513 bait plasmid (“Bait” in the Figure) or with a negative control plasmid (“Vector” in the Figure). The 180 clones obtained in this way were tested again for lacZ activity (not shown) and growth on SD-LWU (central panels) and SD-LWA (right panels) media. Numbers at the left panel correspond to the clone identifiers shown in.+and −: positive and negative controls of protein-protein interaction. Image and information provided by PanBioNet.
(PPTX)
Effects of light intensity on the morphological phenotype of tcu1-1 and tcu1-2. Plants were grown on plates using our standard culture conditions, under continuous light of the photon flux densities shown. Scale bars: 1 mm.
(PPTX)
Arabidopsis mutants used in this work. aAlternative allele names are indicated in parentheses. bSequence obtained in this work. cMolecular nature of the mutation yet to be determined. dBerná et al. (1999). eSIGnAL collection (http://signal.salk.edu). fSAIL collection (http://www.syngenta.com). gParry et al. (2006). hEsteve-Bruna et al. (2013). iLeyser et al. (1993). jRouse et al. (1998).
(DOCX)
Oligonucleotide sets used for the fine mapping of TCU1 . *Labeled with TET (4,7,2′,7′-tetrachloro-6-carboxyfluorescein).
(DOCX)
Other oligonucleotide sets used in this work. a–eThese oligonucleotides include at their 5′ends a EcoRI and b XbaI restriction sites, cthe CACC sequence recognized by the vaccinia virus topoisomerase, and dattB1 and eattB2 sequences, which are shown in italics. fOne half of the primer hybridizes to the 3′ end of one exon and the other half to the 5′ end of the next exon.
(DOCX)
Morphometry of the venation pattern of first- and third-node tcu1-1 leaves. All values are means ± standard deviations from at least 10 measures.
(DOCX)
Results of a Y2H-based screen using TCU1_93-513 as bait.
(DOCX)
Acknowledgments
The authors thank H. Candela for comments on the manuscript, and R. Sarmiento-Mañús, J.M. Serrano, F.M. Lozano, T. Trujillo and L. Serna for technical assistance.
Funding Statement
This research was supported by grants from the Ministerio de Ciencia e Innovación of Spain (BIO2008-01900 to M.R.P.) and the Generalitat Valenciana (PROMETEO/2009/112 to M.R.P. and J.L.M.). A.F.-A. and M.M.A.-P. hold fellowships from the Ministerio de Ciencia e Innovación of Spain. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Bednenko J, Cingolani G, Gerace L (2003) Nucleocytoplasmic transport: Navigating the channel. Traffic 4: 127–135. [DOI] [PubMed] [Google Scholar]
- 2. Sistla S, Vincent PJ, Xia WC, Balasundaram D (2007) Multiple conserved domains of the nucleoporin nup124p and its orthologs Nup1p and Nup153 are critical for nuclear import and activity of the fission yeast Tf1 retrotransposon. Mol Biol Cell 18: 3692–3708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Walde S, Kehlenbach RH (2010) The part and the whole: functions of nucleoporins in nucleocytoplasmic transport. Trends Cell Biol 20: 461–469. [DOI] [PubMed] [Google Scholar]
- 4. Merkle T (2001) Nuclear import and export of proteins in plants: a tool for the regulation of signalling. Planta 213: 499–517. [DOI] [PubMed] [Google Scholar]
- 5. Nakielny S, Dreyfuss G (1999) Transport of proteins and RNAs in and out of the nucleus. Cell 99: 677–690. [DOI] [PubMed] [Google Scholar]
- 6. Rose A, Patel S, Meier I (2004) Plant nuclear envelope proteins. Symp Soc Exp Biol 56: 69–88. [PubMed] [Google Scholar]
- 7. Xu XM, Rose A, Muthuswamy S, Jeong SY, Venkatakrishnan S, et al. (2007) NUCLEAR PORE ANCHOR, the Arabidopsis homolog of Tpr/MIp1/MIp2/megator, is involved in mRNA export and SUMO homeostasis and affects diverse aspects of plant development. Plant Cell 19: 1537–1548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Xu XM, Meulia T, Meier I (2007) Anchorage of plant RanGAP to the nuclear envelope nuclear-pore-associated involves novel proteins. Curr Biol 17: 1157–1163. [DOI] [PubMed] [Google Scholar]
- 9. Zhang YL, Li X (2005) A putative nucleoporin 96 is required for both basal defense and constitutive resistance responses mediated by suppressor of npr1–1, constitutive 1 . Plant Cell 17: 1306–1316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Cheng YT, Germain H, Wiermer M, Bi D, Xu F, et al. (2009) Nuclear pore complex component MOS7/Nup88 is required for innate immunity and nuclear accumulation of defense regulators in Arabidopsis. Plant Cell 21: 2503–2516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Dong CH, Hu X, Tang W, Zheng X, Kim YS, et al. (2006) A putative Arabidopsis nucleoporin, AtNUP160, is critical for RNA export and required for plant tolerance to cold stress. Mol Cell Biol 26: 9533–9543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Cernac A, Lincoln C, Lammer D, Estelle M (1997) The SAR1 gene of Arabidopsis acts downstream of the AXR1 gene in auxin response. Development 124: 1583–1591. [DOI] [PubMed] [Google Scholar]
- 13. Parry G, Ward S, Cernac A, Dharmasiri S, Estelle M (2006) The Arabidopsis SUPPRESSOR OF AUXIN RESISTANCE proteins are nucleoporins with an important role in hormone signaling and development. Plant Cell 18: 1590–1603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Fischer T, Strasser K, Racz A, Rodriguez-Navarro S, Oppizzi M, et al. (2002) The mRNA export machinery requires the novel Sac3p-Thp1p complex to dock at the nucleoplasmic entrance of the nuclear pores. EMBO J 21: 5843–5852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Fischer T, Rodriguez-Navarro S, Pereira G, Racz A, Schiebel E, et al. (2004) Yeast centrin Cdc31 is linked to the nuclear mRNA export machinery. Nat Cell Biol 6: 840–848. [DOI] [PubMed] [Google Scholar]
- 16. Gallardo M, Luna R, Erdjument-Bromage H, Tempst P, Aguilera A (2003) Nab2p and the Thp1p-Sac3p complex functionally interact at the interface between transcription and mRNA metabolism. J Biol Chem 278: 24225–24232. [DOI] [PubMed] [Google Scholar]
- 18. Köhler A, Hurt E (2007) Exporting RNA from the nucleus to the cytoplasm. Nat Rev Mol Cell Biol 8: 761–773. [DOI] [PubMed] [Google Scholar]
- 19. Lu Q, Tang X, Tian G, Wang F, Liu K, et al. (2010) Arabidopsis homolog of the yeast TREX-2 mRNA export complex: components and anchoring nucleoporin. Plant J 61: 259–270. [DOI] [PubMed] [Google Scholar]
- 20. Tamura K, Fukao Y, Iwamoto M, Haraguchi T, Hara-Nishimura I (2010) Identification and characterization of nuclear pore complex components in Arabidopsis thaliana . Plant Cell 22: 4084–4097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Hu TH, Guan TL, Gerace L (1996) Molecular and functional characterization of the p62 complex, an assembly of nuclear pore complex glycoproteins. J Cell Biol 134: 589–601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Berná G, Robles P, Micol JL (1999) A mutational analysis of leaf morphogenesis in Arabidopsis thaliana . Genetics 152: 729–742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana . Science 301: 653–657. [DOI] [PubMed] [Google Scholar]
- 24. Ponce MR, Quesada V, Micol JL (1998) Rapid discrimination of sequences flanking and within T-DNA insertions in the Arabidopsis genome. Plant J 14: 497–501. [DOI] [PubMed] [Google Scholar]
- 25. Mollá-Morales A, Sarmiento-Mañús R, Robles P, Quesada V, Pérez-Pérez JM, et al. (2011) Analysis of ven3 and ven6 reticulate mutants reveals the importance of arginine biosynthesis in Arabidopsis leaf development. Plant J 65: 335–345. [DOI] [PubMed] [Google Scholar]
- 26. Candela H, Martínez-Laborda A, Micol JL (1999) Venation pattern formation in Arabidopsis thaliana vegetative leaves. Dev Biol 205: 205–216. [DOI] [PubMed] [Google Scholar]
- 27. Rubio-Díaz S, Pérez-Pérez JM, González-Bayón R, Muñoz-Viana R, Borrega N, et al. (2012) Cell expansion-mediated organ growth is affected by mutations in three EXIGUA genes. PLoS ONE 7: e36500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. González-Bayón R, Kinsman EA, Quesada V, Vera A, Robles P, et al. (2006) Mutations in the RETICULATA gene dramatically alter internal architecture but have little effect on overall organ shape in Arabidopsis leaves. J Exp Bot 57: 3019–3031. [DOI] [PubMed] [Google Scholar]
- 29. Abramoff MD, Magelhaes PJ, Ram SJ (2004) Image processing with ImageJ. Biophotonics Int 11: 36–42. [Google Scholar]
- 30. Robles P, Fleury D, Candela H, Cnops G, Alonso-Peral MM, et al. (2010) The RON1/FRY1/SAL1 gene is required for leaf morphogenesis and venation patterning in Arabidopsis. Plant Physiol 152: 1357–1372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Ponce MR, Robles P, Lozano FM, Brotons MA, Micol JL (2006) Low-resolution mapping of untagged mutations. Methods Mol Biol 323: 105–113. [DOI] [PubMed] [Google Scholar]
- 32. Ponce MR, Robles P, Micol JL (1999) High-throughput genetic mapping in Arabidopsis thaliana . Mol Gen Genet 261: 408–415. [DOI] [PubMed] [Google Scholar]
- 33. Pérez-Pérez JM, Ponce MR, Micol JL (2004) The ULTRACURVATA2 gene of Arabidopsis encodes an FK506-binding protein involved in auxin and brassinosteroid signaling. Plant Physiol 134: 101–117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Barrero JM, Gonzalez-Bayon R, del Pozo JC, Ponce MR, Micol JL (2007) INCURVATA2 encodes the catalytic subunit of DNA Polymerase alpha and interacts with genes involved in chromatin-mediated cellular memory in Arabidopsis thaliana . Plant Cell 19: 2822–2838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Quesada V, Ponce MR, Micol JL (1999) OTC and AUL1, two convergent and overlapping genes in the nuclear genome of Arabidopsis thaliana . FEBS Lett 461: 101–106. [DOI] [PubMed] [Google Scholar]
- 36. Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana . Plant J 16: 735–743. [DOI] [PubMed] [Google Scholar]
- 37. Quesada V, Macknight R, Dean C, Simpson GG (2003) Autoregulation of FCA pre-mRNA processing controls Arabidopsis flowering time. EMBO J 22: 3142–3152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Heilmann M, Jenkins GI (2013) Rapid reversion from monomer to dimer regenerates the ultraviolet-B photoreceptor UV RESISTANCE LOCUS8 in intact Arabidopsis plants. Plant Physiol 161: 547–555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Zimmermann P, Hirsch-Hoffmann M, Hennig L, Gruissem W (2004) GENEVESTIGATOR. Arabidopsis microarray database and analysis toolbox. Plant Physiol 136: 2621–2632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Peng J, Carol P, Richards DE, King KE, Cowling RJ, et al. (1997) The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses. Genes Dev 11: 3194–3205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Richards DE, King KE, Ait-Ali T, Harberd NP (2001) How gibberellin regulates plant growth and development: a molecular genetic analysis of gibberellin signaling. Annu Rev Plant Physiol Plant Mol Biol 52: 67–88. [DOI] [PubMed] [Google Scholar]
- 42. Silverstone AL, Ciampaglio CN, Sun T (1998) The Arabidopsis RGA gene encodes a transcriptional regulator repressing the gibberellin signal transduction pathway. Plant Cell 10: 155–169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Stamm P, Kumar PP (2010) The phytohormone signal network regulating elongation growth during shade avoidance. J Exp Bot 61: 2889–2903. [DOI] [PubMed] [Google Scholar]
- 44. Kandasamy MK, Gilliland LU, McKinney EC, Meagher RB (2001) One plant actin isovariant, ACT7, is induced by auxin and required for normal callus formation. Plant Cell 13: 1541–1554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Zhao D, Ni W, Feng B, Han T, Petrasek MG, et al. (2003) Members of the Arabidopsis-SKP1-like gene family exhibit a variety of expression patterns and may play diverse roles in Arabidopsis. Plant Physiol 133: 203–217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Stone SL, Callis J (2007) Ubiquitin ligases mediate growth and development by promoting protein death. Curr Opin Plant Biol 10: 624–632. [DOI] [PubMed] [Google Scholar]
- 47. Park MY, Wu G, Gonzalez-Sulser A, Vaucheret H, Poethig RS (2005) Nuclear processing and export of microRNAs in Arabidopsis. Proc Natl Acad Sci USA 102: 3691–3696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Merkle T (2003) Nucleo-cytoplasmic partitioning of proteins in plants: implications for the regulation of environmental and developmental signalling. Curr Genet 44: 231–260. [DOI] [PubMed] [Google Scholar]
- 49. Leyser HM, Pickett FB, Dharmasiri S, Estelle M (1996) Mutations in the AXR3 gene of Arabidopsis result in altered auxin response including ectopic expression from the SAUR-AC1 promoter. Plant J 10: 403–413. [DOI] [PubMed] [Google Scholar]
- 50. Rouse D, Mackay P, Stirnberg P, Estelle M, Leyser O (1998) Changes in auxin response from mutations in an AUX/IAA gene. Science 279: 1371–1373. [DOI] [PubMed] [Google Scholar]
- 51. Serrano-Cartagena J, Robles P, Ponce MR, Micol JL (1999) Genetic analysis of leaf form mutants from the Arabidopsis Information Service collection. Mol Gen Genet 261: 725–739. [DOI] [PubMed] [Google Scholar]
- 52. Serrano-Cartagena J, Candela H, Robles P, Ponce MR, Perez-Perez JM, et al. (2000) Genetic analysis of incurvata mutants reveals three independent genetic operations at work in Arabidopsis leaf morphogenesis. Genetics 156: 1363–1377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Tian Q, Reed JW (1999) Control of auxin-regulated root development by the Arabidopsis thaliana SHY2/IAA3 gene. Development 126: 711–721. [DOI] [PubMed] [Google Scholar]
- 54. Esteve-Bruna D, Perez-Perez JM, Ponce MR, Micol JL (2013) incurvata13, a novel allele of AUXIN RESISTANT6, reveals a specific role for auxin and the SCF complex in Arabidopsis embryogenesis, vascular specification, and leaf flatness. Plant Physiol 161: 1303–1320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. del Pozo JC, Dharmasiri S, Hellmann H, Walker L, Gray WM, et al. (2002) AXR1-ECR1-dependent conjugation of RUB1 to the Arabidopsis Cullin AtCUL1 is required for auxin response. Plant Cell 14: 421–433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Snow CM, Senior A, Gerace L (1987) Monoclonal antibodies identify a group of nuclear pore complex glycoproteins. J Cell Biol 104: 1143–1156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Finlay DR, Meier E, Bradley P, Horecka J, Forbes DJ (1991) A complex of nuclear pore proteins required for pore function. J Cell Biol 114: 169–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Solmaz SR, Chauhan R, Blobel G, Melcak I (2011) Molecular architecture of the transport channel of the nuclear pore complex. Cell 147: 590–602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Melcak I, Hoelz A, Blobel G (2007) Structure of Nup58/45 suggests flexible nuclear pore diameter by intermolecular sliding. Science 315: 1729–1732. [DOI] [PubMed] [Google Scholar]
- 60. Zhao Q, Meier I (2011) Identification and characterization of the Arabidopsis FG-repeat nucleoporin Nup62. Plant Signal Behav 6: 330–334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Meinke D, Muralla R, Sweeney C, Dickerman A (2008) Identifying essential genes in Arabidopsis thaliana . Trends Plant Sci 13: 483–491. [DOI] [PubMed] [Google Scholar]
- 62. Xu XM, Rose A, Meier I (2007) NUA activities at the plant nuclear pore. Plant Signal Behav 2: 553–555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Parry G, Estelle M (2004) Regulation of cullin-based ubiquitin ligases by the Nedd8/RUB ubiquitin-like proteins. Semin Cell Dev Biol 15: 221–229. [DOI] [PubMed] [Google Scholar]
- 64. del Pozo JC, Estelle M (1999) Function of the ubiquitin-proteosome pathway in auxin response. Trends Plant Sci 4: 107–112. [DOI] [PubMed] [Google Scholar]
- 65. Mockaitis K, Estelle M (2008) Auxin receptors and plant development: a new signaling paradigm. Annu Rev Cell Dev Biol 24: 55–80. [DOI] [PubMed] [Google Scholar]
- 66.Casal JJ (2012) Shade avoidance. The Arabidopsis Book.2012/05/15 ed. pp. e0157. [DOI] [PMC free article] [PubMed]
- 67. Alabadí D, Blázquez MA (2009) Molecular interactions between light and hormone signaling to control plant growth. Plant Mol Biol 69: 409–417. [DOI] [PubMed] [Google Scholar]
- 68. Cerdan PD, Chory J (2003) Regulation of flowering time by light quality. Nature 423: 881–885. [DOI] [PubMed] [Google Scholar]
- 69. Devlin PF, Yanovsky MJ, Kay SA (2003) A genomic analysis of the shade avoidance response in Arabidopsis. Plant Physiol 133: 1617–1629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Carabelli M, Possenti M, Sessa G, Ciolfi A, Sassi M, et al. (2007) Canopy shade causes a rapid and transient arrest in leaf development through auxin-induced cytokinin oxidase activity. Genes Dev 21: 1863–1868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Faria AM, Levay A, Wang Y, Kamphorst AO, Rosa ML, et al. (2006) The nucleoporin Nup96 is required for proper expression of interferon-regulated proteins and functions. Immunity 24: 295–304. [DOI] [PubMed] [Google Scholar]
- 72. Wiermer M, Cheng YT, Imkampe J, Li M, Wang D, et al. (2012) Putative members of the Arabidopsis Nup107–160 nuclear pore sub-complex contribute to pathogen defense. Plant J 70: 796–808. [DOI] [PubMed] [Google Scholar]
- 73. Köhler A, Hurt E (2010) Gene regulation by nucleoporins and links to cancer. Mol Cell 38: 6–15. [DOI] [PubMed] [Google Scholar]
- 74. Bermejo R, Kumar A, Foiani M (2012) Preserving the genome by regulating chromatin association with the nuclear envelope. Trends Cell Biol 22: 465–473. [DOI] [PubMed] [Google Scholar]
- 75. Grunwald D, Singer RH (2012) Multiscale dynamics in nucleocytoplasmic transport. Curr Opin Cell Biol 24: 100–106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Capelson M, Liang Y, Schulte R, Mair W, Wagner U, et al. (2010) Chromatin-bound nuclear pore components regulate gene expression in higher eukaryotes. Cell 140: 372–383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Vaquerizas JM, Suyama R, Kind J, Miura K, Luscombe NM, et al. (2010) Nuclear pore proteins nup153 and megator define transcriptionally active regions in the Drosophila genome. PLoS Genet 6: e1000846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Kalverda B, Pickersgill H, Shloma VV, Fornerod M (2010) Nucleoporins directly stimulate expression of developmental and cell-cycle genes inside the nucleoplasm. Cell 140: 360–371. [DOI] [PubMed] [Google Scholar]
- 79. Reeves PH, Murtas G, Dash S, Coupland G (2002) early in short days 4, a mutation in Arabidopsis that causes early flowering and reduces the mRNA abundance of the floral repressor FLC . Development 129: 5349–5361. [DOI] [PubMed] [Google Scholar]
- 80. Murtas G, Reeves PH, Fu YF, Bancroft I, Dean C, et al. (2003) A nuclear protease required for flowering-time regulation in Arabidopsis reduces the abundance of SMALL UBIQUITIN-RELATED MODIFIER conjugates. Plant Cell 15: 2308–2319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Murase K, Hirano Y, Sun TP, Hakoshima T (2008) Gibberellin-induced DELLA recognition by the gibberellin receptor GID1. Nature 456: 459–463. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Leaf cellular phenotypes of tcu1-1 and Ler. (A–D) Adaxial epidermal cells shown as (A, B) interference contrast micrographs and (C, D) diagrams. (E, F) Diagrams of abaxial epidermal cells. Stomata are only partially drawn and appear as circles in C–F. (G, H) Diagrams of cells of the palisade mesophyll subepidermal layer. (I) Boxplot distribution of cell sizes in the tissues and genotypes shown. Boxes are delimited by the first (Q1, lower hinge) and third (Q3, upper hinge) quartiles. Whiskers represent Q1–1.5·IQ (lower) and Q3+1.5·IQ (upper), where IQ = Q3– Q1. ⋄: Mean. –: Median. ○: Extreme maximum outlier (> [Q3+3·IQ]). × : Maximum outlier. Leaves were collected at 21 das (days after stratification). Scale bars: 50 µm.
(PPTX)
Venation pattern in tcu1-1 leaves. Diagrams were drawn from first- and third-node leaves collected 21 das. The leaf margin is shown in orange. Some excisions at the margin were required to flatten tcu1-1 leaves before microscopy. Scale bar: 2 mm.
(PPTX)
Phenotypic complementation of tcu1-1 by the TCU1pro:TCU1 transgene. The plants shown were isolated on medium supplemented with 15 µg·ml−1 hygromicin among the T1 progeny of tcu1-1 plants transformed by infection with Agrobacterium tumefaciens C5851 cells carrying the pGreen0179 plasmid either without any insert (left) or with the TCU1pro:TCU1 insert (right). The picture was taken 26 das. Scale bar: 2 cm.
(PPTX)
Comparison of deduced amino acid sequences of TCU1 and the putative Nup58s of some higher plants. The TCU1/NUP58 protein of Arabidopsis thaliana (At; NP195430.2) is aligned with homologous gene products from Nicotiana tabacum (Nt; ACY30439.1), Vitis vinifera (Vv; XP_002282659), Populus balsamifera subsp. Trichocarpa (Pb; XP_002310761.1), Oryza sativa (Os; NP_001063345.1) and Zea mays (Zm; NP001132589). Amino acid residues identical or similar in all five sequences are shaded black or grey, respectively. The first of the 452 amino acids that are predicted to be missing in the tcu1-1 mutant is shaded red. The alignment was obtained using Clustal X 2.0 (Larkin et al., 2007) and shaded with Boxshade 3.21 (http://www.ch.embnet.org/software/BOX_form.html).
(PPTX)
Expression data output obtained from the Arabidopsis Electronic Fluorescent Pictograph (eFP) Browser for At4g37130 (TCU1) expression levels throughout all Arabidopsis developmental stages.
(PPTX)
Spatial expression analysis of TCU1. GUS staining of TCU1pro:GUS transgenic plants in (A) roots, (B) a cotyledon, (C–I) expanding leaves and whole rosettes. Plant material was collected at the time shown in each picture (in das). Scale bars: (A–C) 0.5 mm and (D–I) 1 mm.
(PPTX)
Confirmation of interactions identified in a Y2H screen with TCU1_93–513 as bait. Yeast PBN204 cells containing three reporters (URA3, lacZ, and ADE2) that are under the control of different GAL promoters were used. Yeast transformants of the TCU1_93–513 bait and 2 different Arabidopsis cDNA AD libraries were spread on SD-LWU (SD without leucine, tryptophan and uracil) selection medium, which supports growth of cells with bait and prey plasmids yielding proteins interacting each other. After selecting yeast colonies on uracil-deficient media, beta-galactosidase activity was monitored. Growth of the URA+ and lacZ+ colonies on adenosine-deficient media was also tested. This three independent reporter system reduces false positives. In order to confirm the interactions found, the prey parts of the plasmids of the positive clones were amplified by PCR and reintroduced into yeast, each with either the TCU1_93–513 bait plasmid (“Bait” in the Figure) or with a negative control plasmid (“Vector” in the Figure). The 180 clones obtained in this way were tested again for lacZ activity (not shown) and growth on SD-LWU (central panels) and SD-LWA (right panels) media. Numbers at the left panel correspond to the clone identifiers shown in.+and −: positive and negative controls of protein-protein interaction. Image and information provided by PanBioNet.
(PPTX)
Effects of light intensity on the morphological phenotype of tcu1-1 and tcu1-2. Plants were grown on plates using our standard culture conditions, under continuous light of the photon flux densities shown. Scale bars: 1 mm.
(PPTX)
Arabidopsis mutants used in this work. aAlternative allele names are indicated in parentheses. bSequence obtained in this work. cMolecular nature of the mutation yet to be determined. dBerná et al. (1999). eSIGnAL collection (http://signal.salk.edu). fSAIL collection (http://www.syngenta.com). gParry et al. (2006). hEsteve-Bruna et al. (2013). iLeyser et al. (1993). jRouse et al. (1998).
(DOCX)
Oligonucleotide sets used for the fine mapping of TCU1 . *Labeled with TET (4,7,2′,7′-tetrachloro-6-carboxyfluorescein).
(DOCX)
Other oligonucleotide sets used in this work. a–eThese oligonucleotides include at their 5′ends a EcoRI and b XbaI restriction sites, cthe CACC sequence recognized by the vaccinia virus topoisomerase, and dattB1 and eattB2 sequences, which are shown in italics. fOne half of the primer hybridizes to the 3′ end of one exon and the other half to the 5′ end of the next exon.
(DOCX)
Morphometry of the venation pattern of first- and third-node tcu1-1 leaves. All values are means ± standard deviations from at least 10 measures.
(DOCX)
Results of a Y2H-based screen using TCU1_93-513 as bait.
(DOCX)