

Abstract
While much effort has focused on detecting positive and negative directional selection in the human genome, relatively little work has been devoted to balancing selection. This lack of attention is likely due to the paucity of sophisticated methods for identifying sites under balancing selection. Here we develop two composite likelihood ratio tests for detecting balancing selection. Using simulations, we show that these methods outperform competing methods under a variety of assumptions and demographic models. We apply the new methods to whole-genome human data, and find a number of previously-identified loci with strong evidence of balancing selection, including several HLA genes. Additionally, we find evidence for many novel candidates, the strongest of which is FANK1, an imprinted gene that suppresses apoptosis, is expressed during meiosis in males, and displays marginal signs of segregation distortion. We hypothesize that balancing selection acts on this locus to stabilize the segregation distortion and negative fitness effects of the distorter allele. Thus, our methods are able to reproduce many previously-hypothesized signals of balancing selection, as well as discover novel interesting candidates.
Author Summary
In the past, balancing selection was a topic of great theoretical interest that received much attention. However, there has been little focus toward developing methods to identify regions of the genome that are under balancing selection. In this article, we present the first set of likelihood-based methods that explicitly model the spatial distribution of polymorphism expected near a site under long-term balancing selection. Simulation results show that our methods outperform commonly-used summary statistics for identifying regions under balancing selection. Finally, we performed a scan for balancing selection in Africans and Europeans using our new methods and identified a gene called FANK1 as our top candidate outside the HLA region. We hypothesize that the maintenance of polymorphism at FANK1 is the result of segregation distortion.
Introduction
Balancing selection maintains variation within a population. Multiple processes can lead to balancing selection. In overdominance, the heterozygous genotype has higher fitness than either of the homozygous genotypes [1], [2]. In frequency-dependent balancing selection, the fitness of an allele is inversely related to its frequency in the population [2], [3]. In a fluctuating or spatially-structured environment, balancing selection can occur when different alleles are favored in different environments over time or geography [2], [4], [5]. Finally, balancing selection can also be a product of opposite directed effects of segregation distortion balanced by negative selection against the distorter [6]. That is, segregation distortion leads to one allele increasing in frequency. However, if that allele is deleterious, then it is reduced in frequency by negative selection. The combined effect of these opposing forces can lead to a balanced polymorphism.
The genetic signatures of long-term balancing selection at a locus can roughly be divided into three categories [2]. The first signature is that the distribution of allele frequencies will be enriched for intermediate frequency alleles. This occurs because the selected locus itself is likely at moderate frequency within the population and, thus, neutral linked loci will also be at intermediate frequency. The second signature is the presence of trans-specific polymorphisms, which are polymorphisms that are shared among species [7]. This is a result of alleles being maintained over long evolutionary time periods, sometimes for millions of years [8]–[10]. The third signature is an increased density of polymorphic sites. This is due to linked neutral loci sharing similar deep genealogies as that of the selected site, increasing the probability of observing mutations at the neutral loci.
The majority of selection scans in humans have focused on positive and negative directional selection. These studies have found evidence of both types of selection, with negative selection being ubiquitous, and the amount and mechanism of positive selection currently being debated [11]–[13]. However, it is unclear how much balancing selection exists in the human genome. Some scans for balancing selection (e.g., Bubb et al.
[14] and Andrés et al.
[15]) have been carried out using summary statistics such as the Hudson-Kreitman-Aguadé (HKA) test [16] and Tajima's 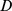 [17] as well as combinations of summary statistics [15], [18] (though see Ségural et al. [7] and Leffler et al.
[19] for recent complementary approaches). The power of such approaches in unclear, and so it is uncertain how important balancing selection is in the human genome. Because balancing selection shapes the genealogy of a sample around a selected locus, more power can be gained by implementing a model of the genealogical process under balancing selection [20], [21]. Composite likelihood methods have proven to be extremely useful for the analysis of genetic variation data using complex population genetic models. [22]–[28]. This approach allows estimation under models without requiring full likelihood calculations, permitting many complex models to be investigated.
[17] as well as combinations of summary statistics [15], [18] (though see Ségural et al. [7] and Leffler et al.
[19] for recent complementary approaches). The power of such approaches in unclear, and so it is uncertain how important balancing selection is in the human genome. Because balancing selection shapes the genealogy of a sample around a selected locus, more power can be gained by implementing a model of the genealogical process under balancing selection [20], [21]. Composite likelihood methods have proven to be extremely useful for the analysis of genetic variation data using complex population genetic models. [22]–[28]. This approach allows estimation under models without requiring full likelihood calculations, permitting many complex models to be investigated.
In this article, we develop two composite likelihood ratio methods to detect balancing selection, which we denote by 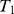 and
and 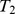 . These methods are based on modeling the effect of balancing selection on the genealogy at linked neutral loci (e.g., Kaplan et al. (1988) [20] and Hudson and Kaplan (1988) [21]) and take into consideration the spatial distributions of polymorphisms and substitutions around a selected site. Through simulations, we show that our methods outperform both HKA and Tajima's
. These methods are based on modeling the effect of balancing selection on the genealogy at linked neutral loci (e.g., Kaplan et al. (1988) [20] and Hudson and Kaplan (1988) [21]) and take into consideration the spatial distributions of polymorphisms and substitutions around a selected site. Through simulations, we show that our methods outperform both HKA and Tajima's 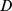 under a variety of demographic assumptions. Further, we apply our methods to autosomal whole-genome sequencing data consisting of nine unrelated European (CEU) and nine unrelated African (YRI) individuals. We find support for multiple targets of balancing selection in the human genome, including previously hypothesized regions such as the human leukocyte antigen (HLA) locus. Additionally, we find evidence for balancing selection at the FANK1 gene, which we hypothesize to result from segregation distortion.
under a variety of demographic assumptions. Further, we apply our methods to autosomal whole-genome sequencing data consisting of nine unrelated European (CEU) and nine unrelated African (YRI) individuals. We find support for multiple targets of balancing selection in the human genome, including previously hypothesized regions such as the human leukocyte antigen (HLA) locus. Additionally, we find evidence for balancing selection at the FANK1 gene, which we hypothesize to result from segregation distortion.
Results
Theory
A new test for balancing selection
In this section, we provide a basic overview of a new test for balancing selection, and we describe the method in greater detail in the sections entitled Kaplan-Darden-Hudson model, Solving the recursion relation, A composite likelihood ratio test based on polymorphism and substitution, and A composite likelihood ratio test based on frequency spectra and substitutions sections. We have developed a new statistical method for detecting balancing selection, which is based on the model of Kaplan, Darden, and Hudson [20], [21] (full details provided in the Kaplan-Darden-Hudson model section). Under this model, we calculate the expected distribution of allele frequencies using simulations, and approximate the probability of observing a fixed difference or polymorphism at a site as a function of its genomic distance to a putative site under balancing selection. Using these calculations, we construct composite likelihood tests that can be used to identify sites under balancing selection, similar to the approaches by Kim and Stephan [23] and Nielsen et al. [26] for detecting selective sweeps.
Basic framework
Consider a biallelic site 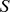 that is under strong balancing selection and maintains an allele
that is under strong balancing selection and maintains an allele 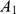 at frequency
at frequency  and an allele
and an allele 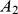 at frequency
at frequency  . Consider a neutral locus
. Consider a neutral locus  that is linked to the selected locus
that is linked to the selected locus 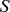 . Denote the scaled recombination rate between the selected locus and the neutral locus as
. Denote the scaled recombination rate between the selected locus and the neutral locus as 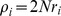 , where
, where 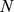 is the diploid population size and
is the diploid population size and 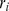 is the per-generation recombination rate. Assume we have a sample of
is the per-generation recombination rate. Assume we have a sample of  genomes from an ingroup species (e.g., humans) and a single genome from an outgroup species (e.g., chimpanzee). From these data, we can estimate the genome-wide expected coalescence time
genomes from an ingroup species (e.g., humans) and a single genome from an outgroup species (e.g., chimpanzee). From these data, we can estimate the genome-wide expected coalescence time 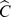 between the ingroup and outgroup species (see Materials and Methods for details). Also, under the Kaplan-Darden-Hudson model, we can obtain the expected tree length
between the ingroup and outgroup species (see Materials and Methods for details). Also, under the Kaplan-Darden-Hudson model, we can obtain the expected tree length 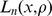 and height
and height 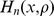 for a sample of
for a sample of  lineages affected by balancing selection by solving a set of recursive equations using the numerical approach described in the Solving the recursion relation. The relationship among
lineages affected by balancing selection by solving a set of recursive equations using the numerical approach described in the Solving the recursion relation. The relationship among 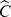 ,
,  , and
, and 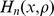 is depicted in Figure 1A
. Assuming a small mutation rate, the probability that a site is polymorphic under a model of balancing selection, given that it contains either a polymorphism or a substitution (fixed difference), is
is depicted in Figure 1A
. Assuming a small mutation rate, the probability that a site is polymorphic under a model of balancing selection, given that it contains either a polymorphism or a substitution (fixed difference), is
| (1) |
and the conditional probability that it contains a substitution is 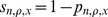 . That is, conditional on a mutation occurring on the genealogy relating the
. That is, conditional on a mutation occurring on the genealogy relating the  ingroup genomes and the outgroup genome, the probability that a site is polymorphic is the probability that a mutation occurs before the most recent common ancestor of the
ingroup genomes and the outgroup genome, the probability that a site is polymorphic is the probability that a mutation occurs before the most recent common ancestor of the  ingroup species (i.e., mutation occurs on red branches indicated in Fig. 1B
), and the probability that a site contains a substitution is the probability that a mutation occurs along the branch leading from the outgroup sequence to the most recent common ancestor of the
ingroup species (i.e., mutation occurs on red branches indicated in Fig. 1B
), and the probability that a site contains a substitution is the probability that a mutation occurs along the branch leading from the outgroup sequence to the most recent common ancestor of the  ingroup species (i.e., mutation occurs on blue branches indicated in Fig. 1C
).
ingroup species (i.e., mutation occurs on blue branches indicated in Fig. 1C
).
Figure 1. Calculation of probabilities of polymorphism and substitution under a model of balancing selection and the incorporation of these probabilities into a genome scan.

(A) Relationship among tree length 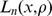 , tree height
, tree height 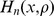 and inter-specific coalescence time
and inter-specific coalescence time 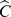 . (B) A site is polymorphic if a mutation occurred on the
. (B) A site is polymorphic if a mutation occurred on the 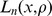 length of branches until the most recent common ancestor of the ingroup sample (red region). (C) A site is a substitution if a mutation occurred on the
length of branches until the most recent common ancestor of the ingroup sample (red region). (C) A site is a substitution if a mutation occurred on the 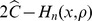 length of branches that represent the divergence between the outgroup species and the most recent common ancestor of the ingroup species (blue region). (D) Height and length of genealogies in relationship to their spatial proximity to a selected site and how the shapes of these genealogies affect the pattern of polymorphism around the site. The composite likelihood ratio is high near a selected site as there is an excess of polymorphisms close to the site and a deficit far from the site.
length of branches that represent the divergence between the outgroup species and the most recent common ancestor of the ingroup species (blue region). (D) Height and length of genealogies in relationship to their spatial proximity to a selected site and how the shapes of these genealogies affect the pattern of polymorphism around the site. The composite likelihood ratio is high near a selected site as there is an excess of polymorphisms close to the site and a deficit far from the site.
Figure 1D shows how the spatial distribution of polymorphism around a selected site is influenced by the underlying genealogy at the site and how this spatial distribution of polymorphism can be used to provide evidence for balancing selection. Within a window of sites, we can obtain the composite likelihood that a particular site is under selection by multiplying the conditional probability of observing a polymorphism or a substitution at every other neutral site as a function of the distance of the neutral site to the balanced polymorphism.
Kaplan-Darden-Hudson model
The genealogy of a neutral locus  linked to the selected locus
linked to the selected locus 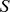 can be traced back in time using the Kaplan, Darden, and Hudson [20], [21] model, which provides a framework for modeling the coalescent process at a neutral locus that is linked to a locus under balancing selection. This model assumes that the selected locus maintains a balanced polymorphism that is infinitely old. Their framework involves modeling selection as a structured population containing two demes representing each of the two allelic classes and migration taking the role of recombination and mutation. Lineages within the first deme are linked to
can be traced back in time using the Kaplan, Darden, and Hudson [20], [21] model, which provides a framework for modeling the coalescent process at a neutral locus that is linked to a locus under balancing selection. This model assumes that the selected locus maintains a balanced polymorphism that is infinitely old. Their framework involves modeling selection as a structured population containing two demes representing each of the two allelic classes and migration taking the role of recombination and mutation. Lineages within the first deme are linked to 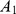 alleles and lineages within the second deme are linked to
alleles and lineages within the second deme are linked to 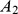 alleles. Lineages migrate between demes by changing their genomic background. That is, a lineage in the first deme will migrate to the second deme if there was a mutation that changed an
alleles. Lineages migrate between demes by changing their genomic background. That is, a lineage in the first deme will migrate to the second deme if there was a mutation that changed an 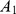 allele to an
allele to an 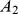 allele or if there was a recombination event that transferred a lineage linked to an
allele or if there was a recombination event that transferred a lineage linked to an 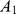 allele to an
allele to an 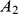 background. Similarly, a lineage in the second deme will migrate to the first deme if there was a mutation that changed an
background. Similarly, a lineage in the second deme will migrate to the first deme if there was a mutation that changed an 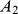 allele to an
allele to an 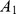 allele or if there was a recombination event that transferred a lineage linked to an
allele or if there was a recombination event that transferred a lineage linked to an 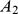 allele to an
allele to an 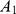 background. The rate at which a lineage linked to an
background. The rate at which a lineage linked to an 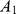 background transfers to an
background transfers to an 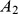 background is
background is 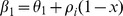 and the rate at which a lineage linked to an
and the rate at which a lineage linked to an 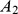 background transfers to an
background transfers to an 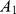 background is
background is 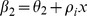 .
.
Consider a sample of  lineages with
lineages with 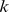 lineages linked to allele
lineages linked to allele 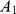 (i.e., in the first deme) and
(i.e., in the first deme) and  lineages linked to allele
lineages linked to allele 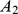 (i.e., in the second deme). Given this configuration, only four events are possible. The first event involves a coalescence of a pair of lineages linked to
(i.e., in the second deme). Given this configuration, only four events are possible. The first event involves a coalescence of a pair of lineages linked to 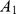 alleles, the second involves a coalescence of a pair of lineages linked to
alleles, the second involves a coalescence of a pair of lineages linked to 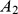 alleles, the third involves the transfer of a lineage from an
alleles, the third involves the transfer of a lineage from an 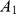 background to an
background to an 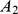 background, and the fourth involves the transfer of a lineage from an
background, and the fourth involves the transfer of a lineage from an 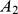 background to an
background to an 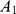 background. The time until the first event (i.e., a coalescence or a transfer of background) is exponentially distributed with rate
background. The time until the first event (i.e., a coalescence or a transfer of background) is exponentially distributed with rate
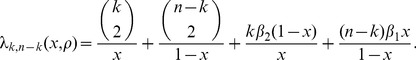 |
(2) |
The probability that the event is a coalescence of a pair of 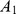 -linked lineages is
-linked lineages is
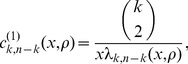 |
(3) |
the event is a coalescence of a pair of 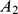 -linked lineages is
-linked lineages is
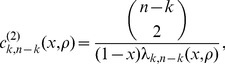 |
(4) |
the event is a transfer from an 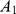 to an
to an 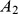 background is
background is
| (5) |
and the event is a transfer from an 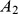 to an
to an 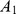 background is
background is
| (6) |
Note that in the notation of Kaplan et al. (1988) [20], 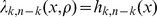 ,
, 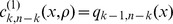 ,
, 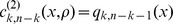 ,
, 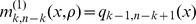 , and
, and 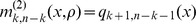 .
.
Let 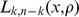 denote the expected tree length given a sample with
denote the expected tree length given a sample with 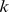
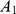 -linked lineages and
-linked lineages and 
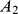 -linked lineages. Using eq. 18 of Kaplan et al. (1988) [20], the expected total tree length can be expressed using the recursion relation
-linked lineages. Using eq. 18 of Kaplan et al. (1988) [20], the expected total tree length can be expressed using the recursion relation
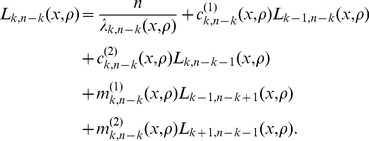 |
(7) |
Similarly, the expected tree height 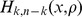 given a sample with
given a sample with 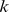
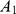 -linked lineages and
-linked lineages and 
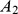 -linked lineages can be expressed by
-linked lineages can be expressed by
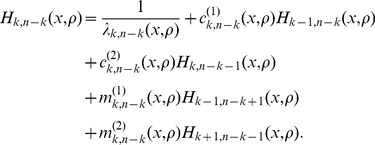 |
(8) |
Solving the recursion relation
Consider a sample of  lineages. Denote the
lineages. Denote the 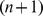 -dimensional vector of tree lengths for a sample of size
-dimensional vector of tree lengths for a sample of size  as
as
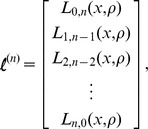 |
such that element  ,
, 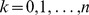 , of
, of 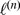 is
is 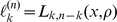 . Next, define the (
. Next, define the (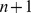 )-dimensional vector
)-dimensional vector
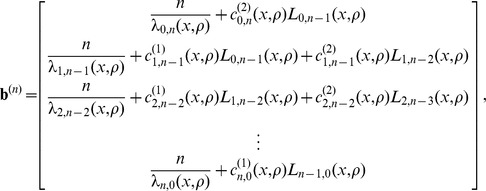 |
such that element 0 is
element  is
is
and element 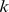 ,
, 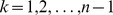 is
is
Further, consider an 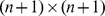 -dimensional tridiagonal matrix of migration rates
-dimensional tridiagonal matrix of migration rates
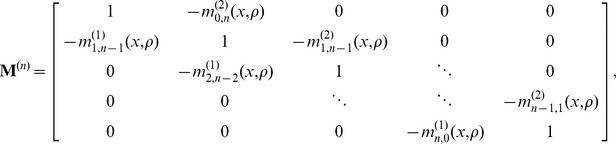 |
with 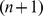 -dimensional main diagonal
-dimensional main diagonal 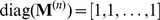 ,
,  -dimensional lower diagonal
-dimensional lower diagonal 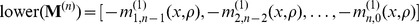 , and
, and  -dimensional upper diagonal
-dimensional upper diagonal 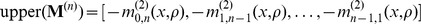 . All elements that do not fall on the main, lower, and upper diagonals of
. All elements that do not fall on the main, lower, and upper diagonals of 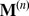 are zero.
are zero.
Given 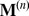 ,
, 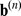 , and
, and 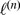 , we can rewrite the recursion relation in eq. 7 as system of equations
, we can rewrite the recursion relation in eq. 7 as system of equations
| (9) |
Because we can calculate eqs. 5 and 6, 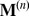 is a constant matrix. For a sample of size
is a constant matrix. For a sample of size  , suppose we know
, suppose we know 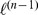 for a sample of size
for a sample of size  . Therefore,
. Therefore, 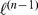 is now a constant vector and hence, because we can calculate eqs. 2–4,
is now a constant vector and hence, because we can calculate eqs. 2–4, 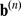 is also a constant vector. Therefore, eq. 9 is a tridiagonal system of
is also a constant vector. Therefore, eq. 9 is a tridiagonal system of 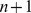 equations with
equations with 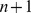 unknowns, which can be solved in
unknowns, which can be solved in 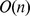 time using the tridiagonal matrix algorithm [29].
time using the tridiagonal matrix algorithm [29].
The base case for the recursion in eq. 8 is when the number of lineages equals one. That is, when all lineages have coalesced and the most recent common ancestor is linked either to an 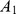 allele or to an
allele or to an 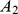 allele. This base case can be represented by
allele. This base case can be represented by 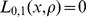 and
and 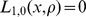 . Given these values, set
. Given these values, set 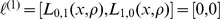 and solve the system of equations
and solve the system of equations 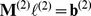 for
for 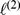 . Next, given
. Next, given 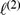 , solve the system of equations
, solve the system of equations 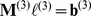 for
for 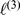 . Iterate this processes until
. Iterate this processes until 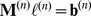 is solved for
is solved for 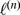 . An analogous process can be used to solve the recursion (eq. 8) for the expected tree height.
. An analogous process can be used to solve the recursion (eq. 8) for the expected tree height.
Using the framework in this section for a sample of size  , we can obtain values for
, we can obtain values for 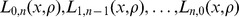 . Given that the
. Given that the 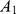 allele has frequency
allele has frequency  and the
and the 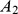 allele has frequency
allele has frequency  , the expected tree length for a sample of size
, the expected tree length for a sample of size  is
is
| (10) |
Similarly, we can obtain the expected tree height 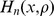 for a sample of size
for a sample of size  . The tree heights and total branch lengths are then used in eq. 1 to compute the likelihood of the data under the selection model.
. The tree heights and total branch lengths are then used in eq. 1 to compute the likelihood of the data under the selection model.
A composite likelihood ratio test based on polymorphism and substitution
In this section, we illustrate how eq. 1 can be incorporated into a composite likelihood. We will then describe a likelihood ratio test that compares the balancing selection model described above to a neutral model based on the background genome patterns of polymorphism. Consider a window of 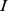 sites that are either polymorphisms or substitutions and consider a putatively selected site
sites that are either polymorphisms or substitutions and consider a putatively selected site 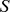 located within the window. Suppose site
located within the window. Suppose site  within the window has
within the window has 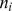 sampled alleles,
sampled alleles, 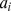 observed ancestral alleles, and is a recombination distance of
observed ancestral alleles, and is a recombination distance of 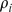 from
from 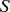 . Let
. Let 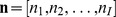 ,
, 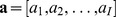 , and
, and 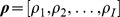 . Define the indicator random variable
. Define the indicator random variable 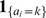 that site
that site  has
has 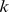 ancestral alleles. Using the Kaplan-Darden-Hudson model, the probability that site
ancestral alleles. Using the Kaplan-Darden-Hudson model, the probability that site  is polymorphic is
is polymorphic is 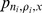 and the probability that the site is a substitution (or fixed difference) is
and the probability that the site is a substitution (or fixed difference) is 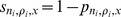 . Under the model, the composite likelihood that site
. Under the model, the composite likelihood that site 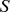 is under balancing selection is
is under balancing selection is
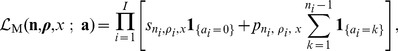 |
(11) |
which is maximized at 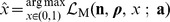 . Notice that sampling distribution for a site depends on the distance to the selected locus. In this method, as in previous composite likelihood methods for detecting selection, there is therefore no need for weighting sites depending on their distance from the selected sites. Such weighting is already incorporated in the probabilistic model. Similarly, there is no need for sliding windows, or the use of Hidden Markov Models (HMMs) to indicate the selected region. The likelihood ratio can, in principle, be calculated for any point in the genome, taking all other points in the genome into account. However, for practical computational reasons, we only calculate the likelihood ratio for a site using nearby sites in a fixed window of 100 substitutions or polymorphisms upstream and downstream of the focal site. As the distance from the selected site increases, little is gained by incorporating information from more sites.
. Notice that sampling distribution for a site depends on the distance to the selected locus. In this method, as in previous composite likelihood methods for detecting selection, there is therefore no need for weighting sites depending on their distance from the selected sites. Such weighting is already incorporated in the probabilistic model. Similarly, there is no need for sliding windows, or the use of Hidden Markov Models (HMMs) to indicate the selected region. The likelihood ratio can, in principle, be calculated for any point in the genome, taking all other points in the genome into account. However, for practical computational reasons, we only calculate the likelihood ratio for a site using nearby sites in a fixed window of 100 substitutions or polymorphisms upstream and downstream of the focal site. As the distance from the selected site increases, little is gained by incorporating information from more sites.
Further, suppose that for a sample of size 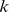 ,
, 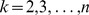 , conditioning only on sites that are polymorphisms or substitutions, the proportion of loci across the genome that are polymorphic is
, conditioning only on sites that are polymorphisms or substitutions, the proportion of loci across the genome that are polymorphic is 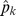 and the proportion of loci that are substitutions is
and the proportion of loci that are substitutions is 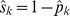 . Then the composite likelihood that site
. Then the composite likelihood that site 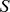 is evolving neutrally is
is evolving neutrally is
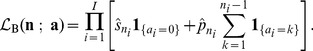 |
(12) |
It follows that the composite likelihood ratio test statistic that site 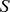 is under balancing selection is
is under balancing selection is 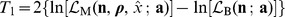 .
.
A composite likelihood ratio test based on frequency spectra and substitutions
A balanced polymorphism not only increases the number of polymorphisms at linked neutral sites, but also leads to an increase in minor allele frequencies at these sites. Therefore, power can be gained by using frequency spectra information in addition to information on the density of polymorphisms and substitutions.
Given a sample of size  , an
, an 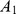 allele at frequency
allele at frequency  ,
, 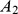 allele at frequency
allele at frequency  , and a polymorphic neutral site that is
, and a polymorphic neutral site that is 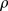 recombination units from a selected site, we can obtain the probability
recombination units from a selected site, we can obtain the probability 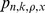 that there are
that there are 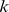 ,
, 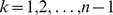 , ancestral alleles observed at the neutral site. The composite likelihood that site
, ancestral alleles observed at the neutral site. The composite likelihood that site 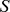 is under balancing selection is
is under balancing selection is
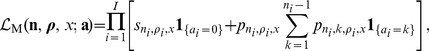 |
(13) |
which is maximized at 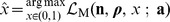 .
.
Further, suppose that for a sample of size 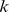 ,
, 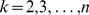 , conditioning only on sites that are polymorphisms or substitutions, the proportion of polymorphic loci across the genome that have
, conditioning only on sites that are polymorphisms or substitutions, the proportion of polymorphic loci across the genome that have 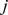 ,
, 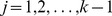 , ancestral alleles is
, ancestral alleles is 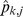 . Then the composite likelihood that site
. Then the composite likelihood that site 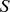 is evolving neutrally is
is evolving neutrally is
| (14) |
It follows that the composite likelihood ratio test statistic that site 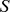 is under balancing selection is
is under balancing selection is 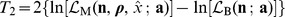 . Because it is computationally difficult to derive analytical formulas for frequency spectra under the Hudson-Darden-Kaplan model, we approximate these distributions by simulating frequency spectra under the Hudson-Darden-Kaplan model for a range of equilibrium frequencies
. Because it is computationally difficult to derive analytical formulas for frequency spectra under the Hudson-Darden-Kaplan model, we approximate these distributions by simulating frequency spectra under the Hudson-Darden-Kaplan model for a range of equilibrium frequencies  and recombination parameters
and recombination parameters 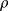 . We then use a look-up table to identify the optimal spectrum to use, and if the optimum is intermediate between two spectra, the two closest distributions are employed. The two new methods,
. We then use a look-up table to identify the optimal spectrum to use, and if the optimum is intermediate between two spectra, the two closest distributions are employed. The two new methods, 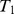 and
and 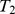 , have been implemented in the software package BALLET (BALancing selection LikElihood Test), which is written in C and is available at http://www.personal.psu.edu/mxd60/software.html.
, have been implemented in the software package BALLET (BALancing selection LikElihood Test), which is written in C and is available at http://www.personal.psu.edu/mxd60/software.html.
Evaluating the methods using simulations
To evaluate the performance of 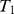 and
and 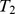 relative to HKA and Tajima's
relative to HKA and Tajima's 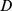 , we carried out extensive simulations of balancing selection using different selection and demographic parameters. We simulated genomic data for a pair of species that diverged
, we carried out extensive simulations of balancing selection using different selection and demographic parameters. We simulated genomic data for a pair of species that diverged 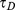 years ago. We introduced a site that is under balancing selection at time
years ago. We introduced a site that is under balancing selection at time 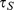 , and the mode of balancing selection at the site is overdominance with selection strength
, and the mode of balancing selection at the site is overdominance with selection strength  and dominance parameter
and dominance parameter 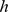 . In the simulations discussed in this article, we varied the demographic history in the target ingroup species, the strength of selection
. In the simulations discussed in this article, we varied the demographic history in the target ingroup species, the strength of selection  , the dominance parameter
, the dominance parameter 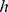 , and the time at which the selected allele arises
, and the time at which the selected allele arises 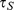 . We consider two values for the strength of selection,
. We consider two values for the strength of selection, 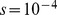 and
and 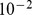 , five values for the dominance parameter,
, five values for the dominance parameter, 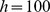 , 10, 3, 1.5, and 1.125, and three times at which the selected allele arises,
, 10, 3, 1.5, and 1.125, and three times at which the selected allele arises, 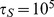 ,
, 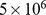 , and
, and 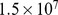 years ago. Under the overdominance model considered here, the equilibrium frequency occurs at
years ago. Under the overdominance model considered here, the equilibrium frequency occurs at 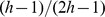 yielding equilibrium frequencies of 0.50, 0.47, 0.40, 0.25, and 0.10 for
yielding equilibrium frequencies of 0.50, 0.47, 0.40, 0.25, and 0.10 for 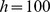 , 10, 3, 1.5, and 1.125, respectively. These parameters were chosen to represent strong (
, 10, 3, 1.5, and 1.125, respectively. These parameters were chosen to represent strong (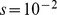 ) and substantially weaker (
) and substantially weaker (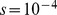 ) selection coefficients and a range of equilibrium frequencies. In addition, the time
) selection coefficients and a range of equilibrium frequencies. In addition, the time 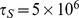 years ago was meant to represent an ancient balanced polymorphism, whereas the other two values for
years ago was meant to represent an ancient balanced polymorphism, whereas the other two values for 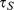 represent violations of assumptions of our methods. That is, the trans-species polymorphism occurring at
represent violations of assumptions of our methods. That is, the trans-species polymorphism occurring at 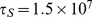 years ago violates the assumption that lineages from the ingroup species are necessarily monophyletic, and the recent balanced polymorphism arising
years ago violates the assumption that lineages from the ingroup species are necessarily monophyletic, and the recent balanced polymorphism arising 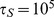 years ago represents balancing selection on an allele that is young relative to the average coalescence time for the ingroup species. Details of how the simulations were implemented are further described in the Materials and Methods section.
years ago represents balancing selection on an allele that is young relative to the average coalescence time for the ingroup species. Details of how the simulations were implemented are further described in the Materials and Methods section.
Ancient balanced polymorphism
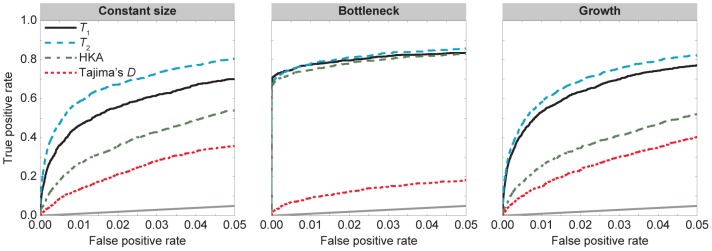
We performed simulations under each of the three demographic models depicted in Figure 2. For these simulations, we constructed receiver operator characteristic (ROC) curves to illustrate relationships between the true and false positive rates of each method. Figure 3 displays ROC curves for 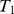 ,
, 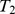 , HKA, and Tajima's
, HKA, and Tajima's 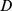 for simulations where
for simulations where 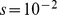 and
and 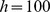 . Under a model of constant population size (left panel of Fig. 3),
. Under a model of constant population size (left panel of Fig. 3), 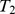 tends to obtain more true positives than
tends to obtain more true positives than 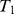 ,
,  more true positives than HKA, and HKA more true positives than Tajima's
more true positives than HKA, and HKA more true positives than Tajima's 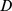 (left panel of Fig. 3). In practice, however, we are typically concerned with a method's performance at low false positive rates. For a false positive rate of
(left panel of Fig. 3). In practice, however, we are typically concerned with a method's performance at low false positive rates. For a false positive rate of 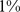 ,
, 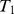 ,
, 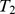 , HKA, and Tajima's
, HKA, and Tajima's 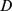 have true positive rates of 30, 40, 14, and
have true positive rates of 30, 40, 14, and 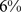 , respectively. Similarly, at a false positive rate of
, respectively. Similarly, at a false positive rate of 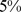 ,
, 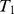 ,
, 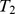 , HKA, and Tajima's
, HKA, and Tajima's 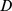 have true positive rates of 58, 67, 37, and
have true positive rates of 58, 67, 37, and 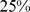 , respectively. These results show that
, respectively. These results show that 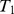 and
and 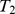 each vastly outperforms both HKA and Tajima's
each vastly outperforms both HKA and Tajima's 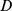 , with
, with 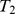 performing better than
performing better than 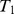 . However, these simulations were performed using the standard neutral model, which is also the demographic model assumed in
. However, these simulations were performed using the standard neutral model, which is also the demographic model assumed in 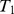 and
and 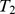 . Thus, to examine the robustness of our methods, we next considered two complex demographic scenarios that could potentially affect the results of our methods—a population bottleneck (Fig. 2B
) and a population expansion (Fig. 2C
).
. Thus, to examine the robustness of our methods, we next considered two complex demographic scenarios that could potentially affect the results of our methods—a population bottleneck (Fig. 2B
) and a population expansion (Fig. 2C
).
Figure 2. Demographic models used in simulations in which a selected allele arises after the split a pair of species.
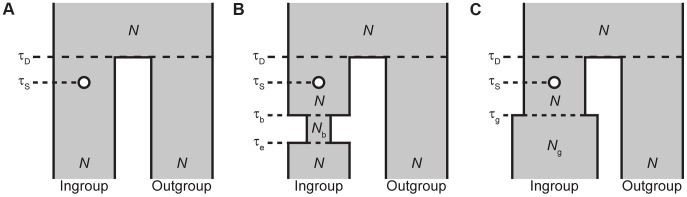
(A) Divergence model. Model parameters are a diploid effective population size 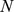 , divergence time
, divergence time 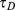 of the ingroup and outgroup species, and the time
of the ingroup and outgroup species, and the time 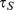 when the selected allele arises. (B) Divergence model with a recent bottleneck within the ingroup species. Additional model parameters are the diploid effective population size
when the selected allele arises. (B) Divergence model with a recent bottleneck within the ingroup species. Additional model parameters are the diploid effective population size 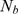 during the bottleneck, the time
during the bottleneck, the time 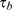 when the bottleneck began, and the time
when the bottleneck began, and the time  when the bottleneck ended. (C) Divergence model with recent population growth within the ingroup species. Additional model parameters are the current diploid effective population size
when the bottleneck ended. (C) Divergence model with recent population growth within the ingroup species. Additional model parameters are the current diploid effective population size 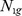 after recent growth and the time
after recent growth and the time 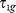 when the growth occurred.
when the growth occurred.
Figure 3. Performance of 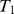 ,
, 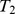 , HKA, and Tajima's
, HKA, and Tajima's 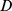 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter  and dominance parameter
and dominance parameter 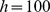 .
.
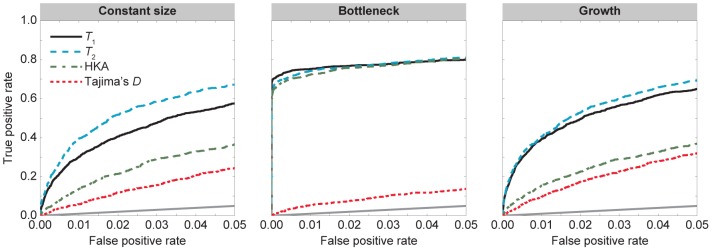
The first column is the divergence model in Figure 2A . The second column is the divergence model in Figure 2B with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C with recent population growth within the ingroup species.
The middle panel of Figure 3 displays ROC curves under a model in which the ingroup species experiences a recent severe bottleneck (Fig. 2B
). For a false positive rate of 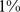 , the true positive rates of
, the true positive rates of 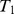 ,
, 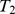 , HKA, and Tajima's
, HKA, and Tajima's 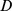 are 75, 74, 72, and
are 75, 74, 72, and 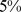 , respectively. Similarly, for a false positive rate of
, respectively. Similarly, for a false positive rate of 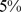 , the true positive rates of
, the true positive rates of 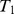 ,
, 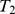 , HKA, and Tajima's
, HKA, and Tajima's 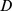 are 80, 81, 80, and
are 80, 81, 80, and 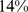 , respectively. Thus, aside from Tajima's
, respectively. Thus, aside from Tajima's 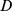 , all methods perform well under this demographic model. This is because a severe population bottleneck decreases levels of diversity across the genome, resulting in a lower polymorphism-to-substitution ratio. Because
, all methods perform well under this demographic model. This is because a severe population bottleneck decreases levels of diversity across the genome, resulting in a lower polymorphism-to-substitution ratio. Because 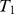 ,
, 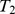 , and HKA all compare levels of polymorphism and divergence at a putatively selected site to those of the corresponding genomic background, these methods are able to identify the increased diversity at a site under balancing selection. In contrast, Tajima's
, and HKA all compare levels of polymorphism and divergence at a putatively selected site to those of the corresponding genomic background, these methods are able to identify the increased diversity at a site under balancing selection. In contrast, Tajima's 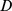 does not perform such a comparison and, thus, has little power to detect balancing selection under this demographic scenario.
does not perform such a comparison and, thus, has little power to detect balancing selection under this demographic scenario.
The right panel of Figure 3 depicts ROC curves under a demographic model in which the ingroup species experiences recent population growth (Fig. 2C
). As with constant population size, 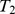 tends to obtain more true positives than
tends to obtain more true positives than 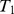 ,
, 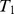 more true positives than HKA, and HKA more true positives than Tajima's
more true positives than HKA, and HKA more true positives than Tajima's 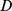 for a given false positive rate. At a false positive rate of
for a given false positive rate. At a false positive rate of 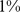 ,
, 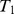 ,
, 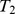 , HKA, and Tajima's
, HKA, and Tajima's 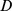 have true positive rates of 39, 41, 15, and
have true positive rates of 39, 41, 15, and 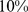 , respectively, and at a false positive rate of
, respectively, and at a false positive rate of 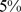 ,
, 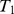 ,
, 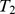 , HKA, and Tajima's
, HKA, and Tajima's 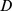 have true positive rates of 65, 69, 37, and
have true positive rates of 65, 69, 37, and 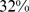 , respectively. Interestingly, all four methods perform better under a recent population growth than under a constant population size. This result is potentially due to less fluctuation in the frequency of a selected allele in the recent past when the population size is large.
, respectively. Interestingly, all four methods perform better under a recent population growth than under a constant population size. This result is potentially due to less fluctuation in the frequency of a selected allele in the recent past when the population size is large.
By considering the demographic models in Figure 2, we have shown that 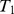 and
and 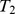 generally outperform both HKA and Tajima's
generally outperform both HKA and Tajima's 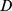 . Next, we investigated the effect of varying
. Next, we investigated the effect of varying 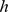 (
(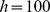 , 10, 3, and 1.5) when
, 10, 3, and 1.5) when 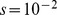 (Fig. S1). Under a model with constant population size (Fig. 2A
),
(Fig. S1). Under a model with constant population size (Fig. 2A
), 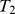 outperforms
outperforms 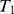 ,
, 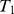 outperforms HKA, and HKA outperforms Tajima's
outperforms HKA, and HKA outperforms Tajima's 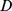 . As
. As 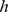 decreases, the performances of HKA and Tajima's
decreases, the performances of HKA and Tajima's 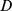 decrease, whereas the performances of
decrease, whereas the performances of 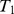 and
and 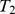 are not dramatically affected. Under a model with a recent population bottleneck (Fig. 2B
),
are not dramatically affected. Under a model with a recent population bottleneck (Fig. 2B
), 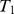 ,
, 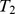 , and HKA all perform well, whereas Tajima's
, and HKA all perform well, whereas Tajima's 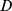 performs poorly. In this scenario,
performs poorly. In this scenario, 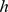 appears to have little influence on the relative performance of these methods. Finally, under a model with a recent population expansion (Fig. 2C
),
appears to have little influence on the relative performance of these methods. Finally, under a model with a recent population expansion (Fig. 2C
), 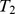 outperforms
outperforms 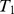 ,
, 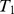 outperforms HKA, and HKA outperforms Tajima's
outperforms HKA, and HKA outperforms Tajima's 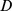 . Decreasing
. Decreasing 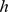 results in a decrease in the performance of Tajima's
results in a decrease in the performance of Tajima's 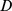 , but has little influence on the performances of all other methods. Moreover, the performances of
, but has little influence on the performances of all other methods. Moreover, the performances of 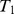 and
and 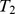 are similar for all
are similar for all 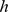 , whereas the perforances of HKA and Tajima's
, whereas the perforances of HKA and Tajima's 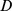 are similar for large
are similar for large 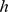 (
(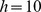 and 100), and dissimilar for low
and 100), and dissimilar for low 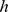 (
(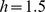 and 3).
and 3).
For 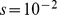 ,
, 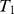 and
and 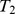 generally perform quite well (Figs. 3 and S1). However, because
generally perform quite well (Figs. 3 and S1). However, because 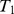 and
and 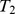 were developed to detect long-term balancing selection of infinite strength, it is unclear how the methods perform under weak selection. To investigate this scenario, we considered
were developed to detect long-term balancing selection of infinite strength, it is unclear how the methods perform under weak selection. To investigate this scenario, we considered 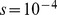 , with
, with 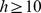 representing relatively strong balancing selection (i.e., relatively high
representing relatively strong balancing selection (i.e., relatively high 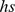 ) and
) and 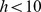 representing relatively weak balancing selection (i.e., relatively low
representing relatively weak balancing selection (i.e., relatively low 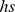 ). For
). For 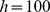 (Fig. 4), we find that the relative performance of the four methods are similar to those in the case of strong selection (
(Fig. 4), we find that the relative performance of the four methods are similar to those in the case of strong selection (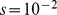 ). Curiously, all methods perform better when
). Curiously, all methods perform better when 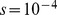 (Fig. 4) than when
(Fig. 4) than when 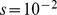 (Fig. 3). To investigate the factors influencing this strange behavior, we plotted the mean difference in the number of polymorphic sites for a scenario with
(Fig. 3). To investigate the factors influencing this strange behavior, we plotted the mean difference in the number of polymorphic sites for a scenario with 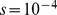 and
and 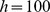 verses one with
verses one with 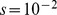 and
and 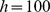 as function of the distance from the site under balancing selection (Fig. S2). We find that, on average, there are more polymorphic sites when the selection coefficient is weak, with the difference in numbers of polymorphic sites disappearing with increasing distance from the site under selection. This phenomenon is due to a drop in local effective population size near the site under balancing selection for the scenario with strong selection. Because
as function of the distance from the site under balancing selection (Fig. S2). We find that, on average, there are more polymorphic sites when the selection coefficient is weak, with the difference in numbers of polymorphic sites disappearing with increasing distance from the site under selection. This phenomenon is due to a drop in local effective population size near the site under balancing selection for the scenario with strong selection. Because 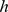 is so large (
is so large (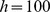 ) and the population size is finite, heterozygous individuals leave a disproportionately large fraction of offspring in the next generation, therefore causing an apparent drop in local effective size near the site under selection.
) and the population size is finite, heterozygous individuals leave a disproportionately large fraction of offspring in the next generation, therefore causing an apparent drop in local effective size near the site under selection.
Figure 4. Performance of 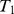 ,
, 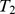 , HKA, and Tajima's
, HKA, and Tajima's 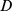 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 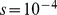 and dominance parameter
and dominance parameter 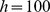 .
.
The first column is the divergence model in Figure 2A . The second column is the divergence model in Figure 2B with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C with recent population growth within the ingroup species.
When 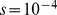 under a model of constant population size (Fig. 2A
),
under a model of constant population size (Fig. 2A
), 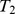 outperforms
outperforms 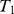 ,
, 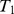 outperforms HKA, and HKA outperforms Tajima's
outperforms HKA, and HKA outperforms Tajima's 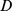 when
when 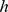 is large (
is large (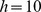 and 100; Fig. S3), similar to what we observe when
and 100; Fig. S3), similar to what we observe when 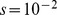 (Fig. S1). In contrast to our observations when
(Fig. S1). In contrast to our observations when 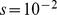 , all methods perform poorly when
, all methods perform poorly when 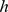 is small (
is small (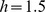 and 3), each identifying signatures of selection only slightly better than random (Fig. S3). Hence, when the selection coefficient is weak and the level of overdominance is low,
and 3), each identifying signatures of selection only slightly better than random (Fig. S3). Hence, when the selection coefficient is weak and the level of overdominance is low, 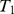 and
and 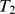 cannot extract enough information from the data to make meaningful predictions. However, HKA and Tajima's
cannot extract enough information from the data to make meaningful predictions. However, HKA and Tajima's 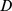 perform just as poorly, and therefore
perform just as poorly, and therefore 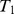 and
and 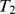 generally outperform HKA and Tajima's
generally outperform HKA and Tajima's 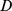 under a demographic model with constant population size.
under a demographic model with constant population size.
Next, when 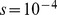 under a model with a recent population bottleneck (Fig. 2B
),
under a model with a recent population bottleneck (Fig. 2B
), 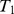 ,
, 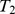 , and HKA all perform well, whereas Tajima's
, and HKA all perform well, whereas Tajima's 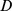 performs poorly (Fig. S3), similar to what we observe when
performs poorly (Fig. S3), similar to what we observe when 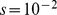 (Fig. S1). In contrast to the results for
(Fig. S1). In contrast to the results for 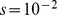 ,
, 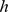 has some influence on the relative performance of these methods. As
has some influence on the relative performance of these methods. As 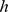 decreases, the performance of all methods decreases—though not substantially. In addition, similarly to what we observe when
decreases, the performance of all methods decreases—though not substantially. In addition, similarly to what we observe when 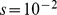 , the performances of
, the performances of 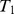 ,
, 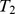 , and HKA are approximately the same. Hence, even under weak selection coefficients, population bottlenecks tend to enhance the performance of
, and HKA are approximately the same. Hence, even under weak selection coefficients, population bottlenecks tend to enhance the performance of 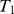 ,
, 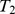 , and HKA, whereas they inhibit the performance of Tajima's
, and HKA, whereas they inhibit the performance of Tajima's 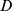 .
.
Finally, when 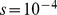 under a model with a recent population expansion (Fig. 2C
),
under a model with a recent population expansion (Fig. 2C
), 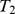 outperforms
outperforms 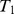 ,
, 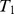 outperforms HKA, and HKA outperforms Tajima's
outperforms HKA, and HKA outperforms Tajima's 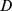 for large
for large 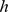 (
(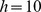 and 100; Fig S3), as observed when
and 100; Fig S3), as observed when 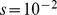 (Fig. S1). In contrast to the results for the case of
(Fig. S1). In contrast to the results for the case of 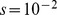 , all methods perform poorly when
, all methods perform poorly when 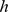 is small (
is small (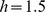 and 3). Hence, like the case under constant population size, when the selection coefficient is weak and the level of overdominance is low,
and 3). Hence, like the case under constant population size, when the selection coefficient is weak and the level of overdominance is low, 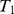 and
and 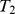 cannot extract enough information from the data to make meaningful predictions. However, HKA and Tajima's
cannot extract enough information from the data to make meaningful predictions. However, HKA and Tajima's 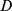 perform just as poorly, and therefore
perform just as poorly, and therefore 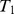 and
and 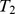 generally outperform HKA and Tajima's
generally outperform HKA and Tajima's 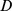 under a demographic model with recent population growth.
under a demographic model with recent population growth.
So far the lowest dominance parameter considered here was 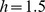 , which has an equilibrium frequency of 0.25. To further assess the limits of our methods, we considered
, which has an equilibrium frequency of 0.25. To further assess the limits of our methods, we considered 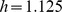 , which has a substantially smaller equilibrium frequency of 0.10. When
, which has a substantially smaller equilibrium frequency of 0.10. When 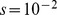 , we find that all four methods perform poorly under the constant population size (Fig. 2A
) and growth (Fig. 2C
) models (Fig. S4). In contrast, as with the higher equilibrium frequencies (Fig. S1),
, we find that all four methods perform poorly under the constant population size (Fig. 2A
) and growth (Fig. 2C
) models (Fig. S4). In contrast, as with the higher equilibrium frequencies (Fig. S1), 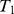 ,
, 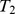 , and HKA statistics performed well, whereas Tajima's
, and HKA statistics performed well, whereas Tajima's 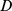 performed poorly under the bottleneck (Fig. 2B
) model (Fig. S4).
performed poorly under the bottleneck (Fig. 2B
) model (Fig. S4).
We next examined violations in recombination rate assumptions of 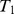 and
and 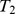 by investigating the robustness of
by investigating the robustness of 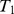 and
and 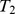 to error in recombination rate estimation. For each simulation, we assumed a recombination rate of
to error in recombination rate estimation. For each simulation, we assumed a recombination rate of 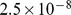 per site per generation. We first wanted to investigate whether using an incorrect recombination map would increase the chances that
per site per generation. We first wanted to investigate whether using an incorrect recombination map would increase the chances that 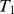 and
and 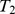 identify false positive. Figure S5 depicts results under a model with constant population size (Fig. 2A
) in which there is no selected allele. With respect to identifying false signals of balancing selection, our results indicate that
identify false positive. Figure S5 depicts results under a model with constant population size (Fig. 2A
) in which there is no selected allele. With respect to identifying false signals of balancing selection, our results indicate that 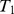 and
and 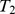 are robust to recombination rate underestimation and overestimation. We next wanted to examine whether using an incorrect recombination map would influence the power of
are robust to recombination rate underestimation and overestimation. We next wanted to examine whether using an incorrect recombination map would influence the power of 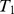 and
and 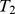 to identify ancient balanced polymorphisms. Figure S6 depicts results for a model with constant population size (Fig. 2A
) with time of selection
to identify ancient balanced polymorphisms. Figure S6 depicts results for a model with constant population size (Fig. 2A
) with time of selection 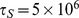 ,
, 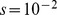 , large (
, large (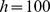 ) and small (
) and small (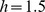 ) dominance parameters, and recombination rate overestimated by one or two orders of magnitude and underestimated by one or two orders of magnitude. We do not consider
) dominance parameters, and recombination rate overestimated by one or two orders of magnitude and underestimated by one or two orders of magnitude. We do not consider 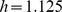 due to the poor performance of all methods considered here for that parameter setting. Incorrectly inferring an order of magnitude higher recombination rate slightly improves the performance of both
due to the poor performance of all methods considered here for that parameter setting. Incorrectly inferring an order of magnitude higher recombination rate slightly improves the performance of both 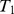 and
and 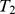 . However, incorrectly inferring a two orders of magnitude higher recombination rate yields poor performance for both
. However, incorrectly inferring a two orders of magnitude higher recombination rate yields poor performance for both 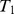 and
and 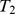 under reasonable false positive rates (e.g., less than 5%). Incorrectly inferring the recombination rate by one or two orders of magnitude lower than the truth does not vastly alter the power for
under reasonable false positive rates (e.g., less than 5%). Incorrectly inferring the recombination rate by one or two orders of magnitude lower than the truth does not vastly alter the power for 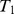 , but substantially decreases the power of
, but substantially decreases the power of 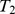 .
.
Ancient trans-species balanced polymorphism
One hallmark of balancing selection is that it maintains polymorphism for a long time, potentially for millions of years [8]–[10]. Thus, some balanced polymorphisms, referred to as trans-specific polymorphisms, are shared across multiple species. Figure S7 displays the three demographic models that we consider in which a selected allele arises in the population ancestral to the split of the ingroup and outgroup species. For each demographic scenario, we set 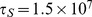 years ago, creating a selected allele that is three times as ancient as the one that we consider in Figure 2. All other model parameters are identical to those considered in Figure 2.
years ago, creating a selected allele that is three times as ancient as the one that we consider in Figure 2. All other model parameters are identical to those considered in Figure 2.
Figures S8 and S9 indicate that the performances of 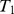 ,
, 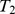 , HKA, and Tajima's
, HKA, and Tajima's 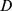 are not greatly affected by considering an ancient trans-species balanced polymorphism when compared to an ancient balanced polymorphism that occurred more recently than the split of a pair of species. This is important because the scenario of an ancient trans-species balanced polymorphism is a violation of the assumptions of the model since it forces lineages from the ingroup species to not be monophyletic with respect to the outgroup species. Hence, though
are not greatly affected by considering an ancient trans-species balanced polymorphism when compared to an ancient balanced polymorphism that occurred more recently than the split of a pair of species. This is important because the scenario of an ancient trans-species balanced polymorphism is a violation of the assumptions of the model since it forces lineages from the ingroup species to not be monophyletic with respect to the outgroup species. Hence, though 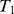 and
and 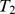 make the assumption that lineages from the ingroup species are monophyletic, this assumption does not hinder the methods in practice.
make the assumption that lineages from the ingroup species are monophyletic, this assumption does not hinder the methods in practice.
Young balanced polymorphism
The two methods developed in this article assume that selection is infinitely strong and that the balanced polymorphism is infinitely old. Here we consider the performance of 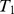 ,
, 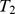 , HKA, and Tajima's
, HKA, and Tajima's 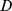 under a scenario in which a young balanced polymorphism arose
under a scenario in which a young balanced polymorphism arose 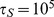 years ago. Considering selection coefficients
years ago. Considering selection coefficients 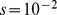 (Fig. S10) and
(Fig. S10) and 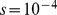 (Fig. S11), all four methods performed poorly under the constant size and growth demographic scenarios, regardless of the dominance parameter. In contrast,
(Fig. S11), all four methods performed poorly under the constant size and growth demographic scenarios, regardless of the dominance parameter. In contrast, 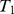 ,
, 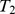 , and HKA all perform well and Tajima's
, and HKA all perform well and Tajima's 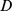 performs poorly under the bottleneck scenario, similar to the results for the ancient balanced polymorphisms. These results show that the new methods have limited power to detect young balanced polymorphisms, except under a scenario in which the background density of polymorphisms is substantially lowered—as in the case of a strong recent population bottleneck.
performs poorly under the bottleneck scenario, similar to the results for the ancient balanced polymorphisms. These results show that the new methods have limited power to detect young balanced polymorphisms, except under a scenario in which the background density of polymorphisms is substantially lowered—as in the case of a strong recent population bottleneck.
Matching the mean density of polymorphisms to a constant size model
The alternate demographic scenarios that we investigated here have focused on the performance of 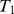 ,
, 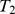 , HKA, and Tajima's
, HKA, and Tajima's 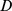 for a recent population bottleneck or growth, relative to a constant size population. However, we have not considered whether a population bottleneck or growth actually changes the absolute performance of the methods, as these demographic events not only change the density of polymorphisms relative to constant size models, but they also change the shape of the frequency spectrum. To control for the density of polymorphisms, we chose the ancestral effective size under the bottleneck and growth models so that the expected number of segregating sites under the bottleneck and growth models is the same as a constant size model of diploid effective size
for a recent population bottleneck or growth, relative to a constant size population. However, we have not considered whether a population bottleneck or growth actually changes the absolute performance of the methods, as these demographic events not only change the density of polymorphisms relative to constant size models, but they also change the shape of the frequency spectrum. To control for the density of polymorphisms, we chose the ancestral effective size under the bottleneck and growth models so that the expected number of segregating sites under the bottleneck and growth models is the same as a constant size model of diploid effective size 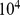 . That is, we set the ancestral sizes for complex demographic models such that these complex models yield identical mean densities of polymorphic sites as a model of constant population size of
. That is, we set the ancestral sizes for complex demographic models such that these complex models yield identical mean densities of polymorphic sites as a model of constant population size of 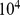 diploid individuals. The details on how we chose these ancestral effective sizes can be found in the Materials and Methods section, with the ancestral diploid effective sizes under the bottleneck and growth models as 14015 and 8762, respectively.
diploid individuals. The details on how we chose these ancestral effective sizes can be found in the Materials and Methods section, with the ancestral diploid effective sizes under the bottleneck and growth models as 14015 and 8762, respectively.
Figures S12 and S13, Figures S14 and S15, and Figures S16 and S17 display results for times 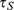 at which a balanced polymorphism arose of
at which a balanced polymorphism arose of 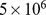 ,
, 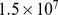 , and
, and 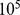 years ago, respectively. Interestingly, these results indicate that the bottleneck and growth models behave similarly to a constant size model once the mean density of polymorphic sites is matched to that of a constant size model. That is, there no longer is a substantial improvement for
years ago, respectively. Interestingly, these results indicate that the bottleneck and growth models behave similarly to a constant size model once the mean density of polymorphic sites is matched to that of a constant size model. That is, there no longer is a substantial improvement for 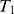 ,
, 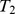 , and HKA for bottleneck models relative to a constant size model. Hence, it is not the shape of the frequency spectrum that gave the apparent increase in power under the bottleneck model (e.g., compare Fig. 3 to Fig. 5 and Fig. 4 to Fig. 6). Rather, it was the large decrease in the background density of polymorphisms relative to that of the assumed effective population size under the model of balancing selection. In addition, when matching the mean density of polymorphisms, methods tended to perform better under the growth model than under the bottleneck model (e.g., Figs. 5 and 6), counter to what was observed without matching the mean density of polymorphisms (e.g., compare Fig. 3 to Fig. 5 and Fig. 4 to Fig. 6). This observation is potentially due to the increased variance in coalescence times under the new bottleneck model compared to the new growth model, when the mean density of polymorphisms is matched to a constant size model.
, and HKA for bottleneck models relative to a constant size model. Hence, it is not the shape of the frequency spectrum that gave the apparent increase in power under the bottleneck model (e.g., compare Fig. 3 to Fig. 5 and Fig. 4 to Fig. 6). Rather, it was the large decrease in the background density of polymorphisms relative to that of the assumed effective population size under the model of balancing selection. In addition, when matching the mean density of polymorphisms, methods tended to perform better under the growth model than under the bottleneck model (e.g., Figs. 5 and 6), counter to what was observed without matching the mean density of polymorphisms (e.g., compare Fig. 3 to Fig. 5 and Fig. 4 to Fig. 6). This observation is potentially due to the increased variance in coalescence times under the new bottleneck model compared to the new growth model, when the mean density of polymorphisms is matched to a constant size model.
Figure 5. Performance of 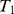 ,
, 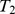 , HKA, and Tajima's
, HKA, and Tajima's 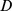 under the bottleneck and growth demographic models in Figure 2 with selection parameter
under the bottleneck and growth demographic models in Figure 2 with selection parameter 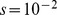 and dominance parameter
and dominance parameter 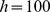 .
.

The left panel is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The right panel is the divergence model in Figure 2C
with recent population growth within the ingroup species. The population sizes for the bottleneck and growth demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size 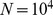 individuals.
individuals.
Figure 6. Performance of 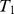 ,
, 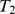 , HKA, and Tajima's
, HKA, and Tajima's 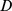 under the bottleneck and growth demographic models in Figure 2 with selection parameter
under the bottleneck and growth demographic models in Figure 2 with selection parameter 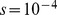 and dominance parameter
and dominance parameter 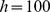 .
.

The left panel is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The right panel is the divergence model in Figure 2C
with recent population growth within the ingroup species. The population sizes for the bottleneck and growth demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size 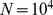 individuals.
individuals.
Empirical analysis
Balancing selection in humans
We probed the effects of balancing selection in humans by using whole-genome sequencing data from nine unrelated individuals from the CEU population and nine unrelated individuals from the YRI population (see Materials and Methods). We performed a scan for balancing selection at each position in our dataset by considering a window of 100 substitutions or polymorphisms upstream and downstream of our focal site. This window size was taken for computational convenience, rather than by consideration of the recombination rate or polymorphism density within the region. Though we used a window size of 200 polymorphisms or substitutions for computational convenience, 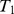 and
and 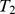 can also be computed using all sites on a chromosome. The mean window length was ∼14.7 kb for the CEU and ∼13.7 kb for the YRI populations, which should be sufficiently long because recombination quickly breaks down the signal of balancing selection at distant neutral sites. That is, under the Hudson-Darden-Kaplan model, the scale at which one would observe an increase in diversity is
can also be computed using all sites on a chromosome. The mean window length was ∼14.7 kb for the CEU and ∼13.7 kb for the YRI populations, which should be sufficiently long because recombination quickly breaks down the signal of balancing selection at distant neutral sites. That is, under the Hudson-Darden-Kaplan model, the scale at which one would observe an increase in diversity is 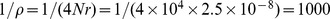 nucleotides, or a 1 kb window [21]. Manhattan plots for
nucleotides, or a 1 kb window [21]. Manhattan plots for 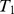 (Figs. S18 and S19) and
(Figs. S18 and S19) and 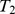 (Figs. S20 and S21) test statistics suggest that there are multiple outlier candidate regions. Intersecting the locations of these scores with those from the longest transcript of each RefSeq gene (i.e., transcription start to stop including exons and introns) led to identification of many previously-hypothesized and novel genes potentially undergoing balancing selection (see Tables S1–S4, with previously-hypothesized genes highlighted in bold).
(Figs. S20 and S21) test statistics suggest that there are multiple outlier candidate regions. Intersecting the locations of these scores with those from the longest transcript of each RefSeq gene (i.e., transcription start to stop including exons and introns) led to identification of many previously-hypothesized and novel genes potentially undergoing balancing selection (see Tables S1–S4, with previously-hypothesized genes highlighted in bold).
Multiple genes at the HLA region are strong outliers (top 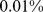 of all scores across the genome) in our scan for balancing selection (Tables S1–S4). Because this study uses high-coverage sequencing data, resolution in the HLA region is particularly fine (Figs. S22 and 7), with strong signals in classical MHC genes such as HLA-A, HLA-B, HLA-C, HLA-DR, HLA-DQ, and HLA-DP genes [14]. The HLA region, which is located on chromosome six, is a well-known site of balancing selection in humans [8]–[10]. The protein products encoded by HLA genes are involved in antigen presentation, thus playing important roles in immune system function. Genes at the HLA locus are known to be highly polymorphic and are thought to be subject to balancing selection due to frequency-dependent selection, overdominance, or fluctuating selection in a rapidly changing pathogenic environment [30], [31]. As the HLA region is so well known as a locus under balancing selection, it is important that our methods identify strong candidate candidate genes in the regions as a proof of concept.
of all scores across the genome) in our scan for balancing selection (Tables S1–S4). Because this study uses high-coverage sequencing data, resolution in the HLA region is particularly fine (Figs. S22 and 7), with strong signals in classical MHC genes such as HLA-A, HLA-B, HLA-C, HLA-DR, HLA-DQ, and HLA-DP genes [14]. The HLA region, which is located on chromosome six, is a well-known site of balancing selection in humans [8]–[10]. The protein products encoded by HLA genes are involved in antigen presentation, thus playing important roles in immune system function. Genes at the HLA locus are known to be highly polymorphic and are thought to be subject to balancing selection due to frequency-dependent selection, overdominance, or fluctuating selection in a rapidly changing pathogenic environment [30], [31]. As the HLA region is so well known as a locus under balancing selection, it is important that our methods identify strong candidate candidate genes in the regions as a proof of concept.
Figure 7. Signals of balancing selection within the HLA region for the CEU (blue) and YRI (orange) populations using the 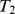 test statistic.
test statistic.

From bottom to top, the horizontal dotted gray lines indicate the 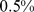 ,
, 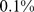 ,
, 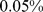 , and
, and 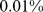 empirical cutoffs, respectively.
empirical cutoffs, respectively.
One gene that we found particularly intriguing is FANK1 (Figs. S23 and 8). This gene is one of the top four candidates in the CEU and YRI populations when using either the 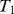 or
or 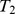 statistic (Tables S1–S4). In addition, FANK1 is the top candidate among genes that have not been previously hypothesized to be under balancing selection when using either test in the CEU and the
statistic (Tables S1–S4). In addition, FANK1 is the top candidate among genes that have not been previously hypothesized to be under balancing selection when using either test in the CEU and the 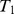 test in the YRI. FANK1 is expressed during the transition from diploid to haploid state in meiosis [32], [33]. Though it is often identified as spermatogenesis-specific [32], [33], it is also expressed during oogenesis in cattle [34] and mice [35]. Its function is to suppress apoptosis [33], and it is one of ten to 20 genes identified as being imprinted in humans (i.e., allele specific methylation) [36]. Interestingly, it also shows marginal evidence of segregation distortion (Fig. 8) [37]. Further, as a CpG island resides directly underneath our signal in both the CEU and YRI populations, we analyzed the region around FANK1 with all
test in the YRI. FANK1 is expressed during the transition from diploid to haploid state in meiosis [32], [33]. Though it is often identified as spermatogenesis-specific [32], [33], it is also expressed during oogenesis in cattle [34] and mice [35]. Its function is to suppress apoptosis [33], and it is one of ten to 20 genes identified as being imprinted in humans (i.e., allele specific methylation) [36]. Interestingly, it also shows marginal evidence of segregation distortion (Fig. 8) [37]. Further, as a CpG island resides directly underneath our signal in both the CEU and YRI populations, we analyzed the region around FANK1 with all 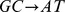 transitions on chromosome 10 removed as well as all transitions on chromosome 10 removed and we still retain the peak (Fig. S24), strongly suggesting that the signature of balancing selection that we identified around FANK1 is not driven by CpG mutational effects. We were additionally surprised to find that the putative selection signal was approximately 40 kb wide, which is abnormally large for balancing selection. Looking back at the recombination map, we find that the rates in this region are extremely low, which explains the large width of the peak. However, Figures S5 and S6 indicate that erroneously inferring a lower recombination rate does not increase the power of detecting a selection signal, and can substantially impair the ability for
transitions on chromosome 10 removed as well as all transitions on chromosome 10 removed and we still retain the peak (Fig. S24), strongly suggesting that the signature of balancing selection that we identified around FANK1 is not driven by CpG mutational effects. We were additionally surprised to find that the putative selection signal was approximately 40 kb wide, which is abnormally large for balancing selection. Looking back at the recombination map, we find that the rates in this region are extremely low, which explains the large width of the peak. However, Figures S5 and S6 indicate that erroneously inferring a lower recombination rate does not increase the power of detecting a selection signal, and can substantially impair the ability for 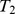 to detect a selection signal.
to detect a selection signal.
Figure 8. Signal of balancing selection at the FANK1 gene for the CEU (blue) and YRI (orange) populations using the 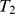 test statistic.
test statistic.

From bottom to top, the horizontal dotted gray lines indicate the 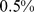 ,
, 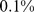 ,
, 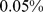 , and
, and 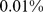 empirical cutoffs, respectively. SNPs (rsIDs) correspond to markers showing significant levels of transmission distortion within the Meyer et al. study [37].
empirical cutoffs, respectively. SNPs (rsIDs) correspond to markers showing significant levels of transmission distortion within the Meyer et al. study [37].
More broadly, a glance at the top signals for the CEU (Tables S1 and S3) and YRI (Tables S2 and S4) populations, reveals a substantial overlap in the candidate genes identified between the pair. If balancing selection has maintained a polymorphism for a long period of time, then we would expect these populations to share many signals in common due to their relatively recent population split. Tables S1–S4 indicate that our scan also identified a number of genes that were previously-hypothesized to be under balancing selection. However, the majority of this overlap is due to the HLA region. One candidate that we did not find support for was the ABO gene, which has been identified as a potential strong candidate using diverse complementary approaches such as summary statistics [38] and trans-specific polymorphism information [7]. A number of factors, including the small sample size for each of the CEU and YRI populations used here and potential differences in the Complete Genomics dataset relative to others, could have caused the ABO gene to not be at the top of our list of candidates.
Gene ontology analysis
To elucidate functional similarities among genes identified to be under balancing selection, we performed gene ontology (GO) enrichment analysis using GOrilla
[39], [40]. First, we compared an unranked list of the top 100 candidate genes (Tables S1–S4) to the background list of all unique genes. Genes obtained using either test statistic are enriched for processes involved in the immune response in both the CEU and YRI populations (Tables S5–S8). Similarly, the top genes are enriched for MHC class II functional categories (Tables S9–S11), with the exception of the 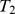 statistic applied to YRI, which has no functional enrichment. Further, these top genes tend to be components of the MHC complex and membranes (Tables S12–S15), which often directly interact with pathogens. Interestingly, removing all HLA genes from both the top 100 and background sets of genes reveals no GO enrichment for process, function, or component categories, indicating that enrichment is predominately driven by the HLA region. Because we can also provide a score for each candidate gene in our likelihood framework, we performed a second analysis in which we ranked genes by their likelihood ratio test statistic, with the goal of identifying GO categories that are enriched in top-ranked genes. Using this framework, the top candidate genes tend to be involved in immune response and cell adhesion processes (Tables S16–S19); MHC activity and membrane protein activity functions, such as transporting and binding molecules (Tables S20–S23); and MHC complex, membrane, and cell junction components (Tables S24–S27). In contrast to the case of the top 100 candidate genes, removing all HLA genes from the ranked list still resulted in GO enrichment in categories such as cell adhesion (processes), membrane protein activity (function), and components of membranes and cell junctions (component).
statistic applied to YRI, which has no functional enrichment. Further, these top genes tend to be components of the MHC complex and membranes (Tables S12–S15), which often directly interact with pathogens. Interestingly, removing all HLA genes from both the top 100 and background sets of genes reveals no GO enrichment for process, function, or component categories, indicating that enrichment is predominately driven by the HLA region. Because we can also provide a score for each candidate gene in our likelihood framework, we performed a second analysis in which we ranked genes by their likelihood ratio test statistic, with the goal of identifying GO categories that are enriched in top-ranked genes. Using this framework, the top candidate genes tend to be involved in immune response and cell adhesion processes (Tables S16–S19); MHC activity and membrane protein activity functions, such as transporting and binding molecules (Tables S20–S23); and MHC complex, membrane, and cell junction components (Tables S24–S27). In contrast to the case of the top 100 candidate genes, removing all HLA genes from the ranked list still resulted in GO enrichment in categories such as cell adhesion (processes), membrane protein activity (function), and components of membranes and cell junctions (component).
Discussion
In this article, we presented two likelihood-based methods, 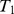 and
and 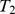 , to identify genomic sites under balancing selection. These methods combine intra-species polymorphism and inter-species divergence with the spatial distribution of polymorphisms and substitutions around a selected site. Through simulations, we showed that
, to identify genomic sites under balancing selection. These methods combine intra-species polymorphism and inter-species divergence with the spatial distribution of polymorphisms and substitutions around a selected site. Through simulations, we showed that 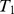 and
and 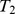 vastly outperform both the HKA test and Tajima's
vastly outperform both the HKA test and Tajima's 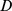 under a diverse set of demographic assumptions, such as a population bottleneck and growth. In addition, application of
under a diverse set of demographic assumptions, such as a population bottleneck and growth. In addition, application of 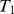 and
and 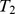 to whole-genome sequencing data from Europeans and Africans revealed many previously identified and novel loci displaying signatures of balancing selection.
to whole-genome sequencing data from Europeans and Africans revealed many previously identified and novel loci displaying signatures of balancing selection.
Simulation results suggest that 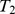 performs at least as well as
performs at least as well as 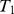 , and so a natural question is whether
, and so a natural question is whether 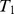 would ever be used. Based on the fact that
would ever be used. Based on the fact that 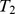 uses the allele frequency spectrum and
uses the allele frequency spectrum and 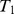 does not, then
does not, then 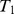 would be a valuable statistic to employ when allele frequencies cannot be estimated well. One example is a situation in which the sample size is small (e.g., one or two genomes). Under this scenario, the
would be a valuable statistic to employ when allele frequencies cannot be estimated well. One example is a situation in which the sample size is small (e.g., one or two genomes). Under this scenario, the 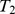 test statistic would likely provide little additional power over the
test statistic would likely provide little additional power over the 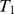 statistic. As another example, it is becoming increasingly common for studies to sequence a pooled sample of individuals rather than each individual in the sample separately. This pooled sequencing will tend to yield inaccurate estimates of allele frequencies across the genome, which could heavily influence the performance of the
statistic. As another example, it is becoming increasingly common for studies to sequence a pooled sample of individuals rather than each individual in the sample separately. This pooled sequencing will tend to yield inaccurate estimates of allele frequencies across the genome, which could heavily influence the performance of the 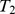 statistic. However, if there is sufficient enough evidence that a site has a pair of alleles observed in the sample, then this site can be considered polymorphic regardless of its actual allele frequency. Future developments that can statistically account for this uncertainty in allele frequency estimation could be incorporated into the
statistic. However, if there is sufficient enough evidence that a site has a pair of alleles observed in the sample, then this site can be considered polymorphic regardless of its actual allele frequency. Future developments that can statistically account for this uncertainty in allele frequency estimation could be incorporated into the 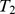 test statistic so that it can be applied to pooled sequencing data. In addition, our investigation into the robustness of
test statistic so that it can be applied to pooled sequencing data. In addition, our investigation into the robustness of  and
and 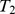 to errors in recombination rate estimates suggested that
to errors in recombination rate estimates suggested that 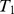 tends to perform better than
tends to perform better than 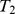 when the estimate of the recombination rate is inaccurate. Because reliable genetic maps are unavailable for most organisms that have had their genome sequenced,
when the estimate of the recombination rate is inaccurate. Because reliable genetic maps are unavailable for most organisms that have had their genome sequenced, 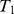 may be the preferable statistic for many current applications.
may be the preferable statistic for many current applications.
The model of balancing selection used in this article is from Hudson and Kaplan [21], and assumes that natural selection is so strong that it maintains a constant allele frequency at the selected locus forever. The simulation scenarios considered here assumed that the strength of balancing selection was also constant since the selected allele arose. However, selection coefficients can fluctuate over time, which provides the basis for future work on investigating the robustness of methods for detecting balancing selection under scenarios in which the strength of selection fluctuates or when selection is weak. Future work can use the framework developed here to construct methods for identifying balancing selection under models with more relaxed assumptions (e.g., see Barton and Etheridge [41] and Barton et al. [42] for potential models).
Recall that we chose a window size based on a fixed number of polymorphisms and substitutions. However, we could have chosen a window in a different way. For example, a window could have been chosen based on physical or genetic distance, rather than a fixed number of substitutions or polymorphisms. However, basing each likelihood calculation on a fixed number of substitutions or polymorphisms, rather than physical or genetic distance, enables each likelihood ratio to be based on the same number of terms, thereby letting the likelihood ratio depend on the density of polymorphisms vs. substitutions rather than the number of polymorphisms in the window. This contrasts other composite likelihood approaches for detecting positive selection (e.g., Nielsen et al., 2005 [26]), where the likelihood under the selection model approaches the likelihood under neutrality with increasing distance from the site under selection. This characteristic exhibited by these other composite likelihood approaches permits variable-size windows, so that at some point adding new terms to the likelihood ratio will not change its value. However, for our method, the likelihood under selection does not approach the likelihood under the background level of diversity (neutrality) with increasing distance from the putative site under selection, causing the value of the likelihood ratio to change by modifying the number of terms. If we chose a standard neutral model for the null hypothesis, then the likelihood under selection would approach the likelihood under the null model with increasing distance from the selected site. To attempt to account for demographic history, we have instead chosen to use the genome-wide level of diversity for the null hypothesis, which does not require that the likelihood under selection to approach the likelihood under the null hypothesis with increasing distance from the putative balanced polymorphism.
In our empirical analysis, we calculated the likelihood ratio (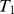 or
or 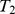 ) for numerous positions along the genome. We then ranked genes according to the largest likelihood ratio estimated between the annotated transcription start and stop of the gene. A consequence of ranking genes in this manner is that longer genes are more likely to be significant. However, because ancient balancing selection only impacts a relatively small region of the genome (in contrast to recent positive selection), the signal of ancient balancing selection could be masked if we instead assigned the average likelihood ratio as the score for a large gene. We therefore opted to assign the score for a gene as the highest likelihood ratio calculated within that gene.
) for numerous positions along the genome. We then ranked genes according to the largest likelihood ratio estimated between the annotated transcription start and stop of the gene. A consequence of ranking genes in this manner is that longer genes are more likely to be significant. However, because ancient balancing selection only impacts a relatively small region of the genome (in contrast to recent positive selection), the signal of ancient balancing selection could be masked if we instead assigned the average likelihood ratio as the score for a large gene. We therefore opted to assign the score for a gene as the highest likelihood ratio calculated within that gene.
Our methods have been shown to be substantially more powerful than HKA and Tajima's 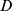 at detecting ancient balanced polymorphisms. However, a glance at Figures 3 and 4 indicates that under constant size and growth models our methods have little power to detect balanced polymorphisms at low false positive rates—a range that would be necessary to detect ancient balancing selection if it were rare. Hence, if balancing selection is relatively rare, then relying solely on statistics considered here to identify ancient balanced polymorphisms could possibly lead to an overabundance of false positives. Complementary evidence, such as considering patterns of linkage disequilibrium or trans-specific polymorphisms in candidate regions, should also be employed to hone in on true signals of ancient balancing selection.
at detecting ancient balanced polymorphisms. However, a glance at Figures 3 and 4 indicates that under constant size and growth models our methods have little power to detect balanced polymorphisms at low false positive rates—a range that would be necessary to detect ancient balancing selection if it were rare. Hence, if balancing selection is relatively rare, then relying solely on statistics considered here to identify ancient balanced polymorphisms could possibly lead to an overabundance of false positives. Complementary evidence, such as considering patterns of linkage disequilibrium or trans-specific polymorphisms in candidate regions, should also be employed to hone in on true signals of ancient balancing selection.
Though we have shown that 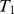 and
and 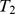 perform well under a population bottleneck and growth, they may be less robust to other forms of demographic model violations, such as population structure. Because population subdivision increases the time to coalescence and corresponding length of a genealogy, we expect higher levels of polymorphism across the genome. Under most assumptions, population subdivision affects the genome uniformly; it increases the level of background polymorphism and likely only slightly decreases the power of the new statistics. However, in some cases, such as an ancient admixture event (e.g., with Neanderthals [43] or Denisovans [44]), levels of variability may increase in only a few regions of the genome, increasing the mean coalescence time in these regions. Such regions may appear to have excess polymorphism relative to background levels and, hence, display false signals of balancing selection under the
perform well under a population bottleneck and growth, they may be less robust to other forms of demographic model violations, such as population structure. Because population subdivision increases the time to coalescence and corresponding length of a genealogy, we expect higher levels of polymorphism across the genome. Under most assumptions, population subdivision affects the genome uniformly; it increases the level of background polymorphism and likely only slightly decreases the power of the new statistics. However, in some cases, such as an ancient admixture event (e.g., with Neanderthals [43] or Denisovans [44]), levels of variability may increase in only a few regions of the genome, increasing the mean coalescence time in these regions. Such regions may appear to have excess polymorphism relative to background levels and, hence, display false signals of balancing selection under the 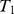 statistic. However, in non-African humans, introgressed regions typically have low population frequencies [43], [44], and, hence, it would be unlikely for polymorphic sites in these regions to harbor many introgressed alleles segregating at intermediate frequencies. Thus, the
statistic. However, in non-African humans, introgressed regions typically have low population frequencies [43], [44], and, hence, it would be unlikely for polymorphic sites in these regions to harbor many introgressed alleles segregating at intermediate frequencies. Thus, the 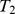 statistic, which explicitly utilizes allele frequency spectra information, would likely be able to distinguish these blocks of archaic admixture from regions of balancing selection. Further, as observed in other studies of natural selection [45], [46], increased robustness to confounding demographic processes can potentially be gained through the use of additional information. For example, population bottlenecks as well as gene flow can increase linkage disequilibrium [47], [48]. Therefore, knowledge about linkage disequilibrium in a region could aid in distinguishing population subdivision from long-term balancing selection.
statistic, which explicitly utilizes allele frequency spectra information, would likely be able to distinguish these blocks of archaic admixture from regions of balancing selection. Further, as observed in other studies of natural selection [45], [46], increased robustness to confounding demographic processes can potentially be gained through the use of additional information. For example, population bottlenecks as well as gene flow can increase linkage disequilibrium [47], [48]. Therefore, knowledge about linkage disequilibrium in a region could aid in distinguishing population subdivision from long-term balancing selection.
Another concern when performing genomes scans for balancing selection is the possibility of false positives due to bioinformatical errors. For example, misalignment of sequence reads in duplicated regions may lead to falsely elevated levels of variability. In many cases, this problem can be alleviated by removing duplicated regions from analyses. However, a non-negligible portion of the human genome is not represented in standard reference sequences and, thus, there may be many unidentified paralogs in the genome. Fortunately, removing sites that deviate from Hardy-Weinberg equilibrium helps to alleviate these problems, because SNPs fixed between or segregating at high frequencies in one of two (or more) paralogous regions will have an excess of heterozygotes in combined short-read alignments. We applied a Hardy-Weinberg filter to all empirical data analyzed in this article. We note that deviations from Hardy-Weinberg equilibrium are expected under certain forms of balancing selection. In theory, a balancing selection signal could, therefore, be lost due to such filtering. However, we used a filtering cutoff of 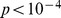 (see Materials and Methods). The strength of selection required to cause this type of deviation from Hardy-Weinberg equilibrium used in the filtering is extremely strong, and such selection would almost certainly have been detected using other methods. Well-established examples of balancing selection in the human genome, such as the selection affecting the HLA loci, are not lost because of filtering, and would generally not be easily detectable using deviations from Hardy-Weinberg as a test. Nonetheless, because phenomena other than balancing selection, such as bioinformatical errors or archaic admixture, could potentially lead to false signals of balancing selection, additional evidence should be obtained before definitively concluding that a site has been subjected to balancing selection.
(see Materials and Methods). The strength of selection required to cause this type of deviation from Hardy-Weinberg equilibrium used in the filtering is extremely strong, and such selection would almost certainly have been detected using other methods. Well-established examples of balancing selection in the human genome, such as the selection affecting the HLA loci, are not lost because of filtering, and would generally not be easily detectable using deviations from Hardy-Weinberg as a test. Nonetheless, because phenomena other than balancing selection, such as bioinformatical errors or archaic admixture, could potentially lead to false signals of balancing selection, additional evidence should be obtained before definitively concluding that a site has been subjected to balancing selection.
One source of additional evidence of balancing selection is whether a signal lies within a region harboring a trans-specific polymorphism [7], [19] because it is unlikely to have a polymorphism segregating in each of a pair of closely-related species without selection maintaining the polymorphism. However, relying solely on evidence from trans-specific polymorphisms would miss many true signals of balancing selection that are not maintained as trans-specific polymorphisms. In addition, regions with bioinformatical errors (e.g., mapping errors) may give the same errors in both species, resulting in a false signal of a shared polymorphism between the pair of species. Nevertheless, the observation of a trans-specific polymorphism can provide convincing evidence of an ancient balanced polymorphism [7], [19]. Previous studies of selection have shown that combinations of statistics can be powerful tools when identifying genes under selection [15], [18], [49]. Hence, combining our methods with other summaries (e.g., linkage disequilibrium [45]–[48]) or information on trans-species polymorphisms [7], [19] will lead to increasingly effective approaches for detecting balancing selection.
The current approach taken by 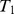 and
and 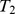 ignores higher order linkage disequilibria, in the sense that it ignores linkage disequilibrium between pairs of neutral markers and only considers correlations between neutral markers and the site under selection. However, incorporating higher order linkage information, such as employing tests based on haplotypes, could provide some advantage. For example,
ignores higher order linkage disequilibria, in the sense that it ignores linkage disequilibrium between pairs of neutral markers and only considers correlations between neutral markers and the site under selection. However, incorporating higher order linkage information, such as employing tests based on haplotypes, could provide some advantage. For example, 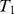 and
and 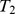 have little power to detect young balanced polymorphisms. However, the haplotype pattern around a young balanced polymorphism is likely to mimic that of an incomplete or partial selective sweep. Therefore, methods that use haplotype information (e.g., EHH [50], iHS [51], and
have little power to detect young balanced polymorphisms. However, the haplotype pattern around a young balanced polymorphism is likely to mimic that of an incomplete or partial selective sweep. Therefore, methods that use haplotype information (e.g., EHH [50], iHS [51], and 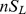 [52]), could provide a complementary and powerful approach to detecting recent balancing selection—a selective regime that the methods considered here have little power.
[52]), could provide a complementary and powerful approach to detecting recent balancing selection—a selective regime that the methods considered here have little power.
Another commonly-cited source of evidence for balancing selection is based on consideration of the topology and branch lengths of within-species haplotype trees. Under long-term balancing selection, the underlying genealogy (e.g., see Fig. S25) will be symmetric, with long basal branches separating a pair of allelic classes (i.e., haplotypes containing one variant and haplotypes containing the other variant). However, the underlying genealogy for a linked neutral variant may differ substantially from that of the selected site. Around a balanced polymorphism, there will be a strong reduction of linkage disequilibrium, not unlike a recombination hotspot, because the long genealogy in the balanced polymorphism provides extra opportunities for recombination. Consequently, the signal of balancing selection will be narrow, and trees estimated from sites located in a window around the balanced polymorphism may fail to detect the presence of highly divergent haplotypes. The utility of within-species haplotype trees as a signature of long-term balancing selection is unclear, as the genealogy of the haplotype may not match the genealogy of the selected region. For example, Figure S26 shows that haplotype trees based on scenarios under balancing selection appear similar to those under neutrality, with the difference that external branches are slightly longer under balancing selection than under neutrality, which contrasts with the generally-held belief that basal branches should be long. These inferred long external branches are a product of estimating haplotype trees in recombining regions [53], which would likely be unavoidable in genomic regions under ancient balancing selection even if recombination events were undetected. As such, haplotype networks or trees built without explicitly accounting for recombination may not be powerful tools for identifying regions under balancing selection.
An assumption of the methods 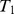 and
and 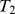 introduced in this article is that two allelic classes at a selected site are maintained for an infinitely long period of time at a constant equilibrium frequency by balancing selection. However, balancing selection is not restricted to act only on two stable allelic classes, and the equilibrium frequency can fluctuate with time and space. Examples of balancing selection that do not conform to our model assumptions are frequency-dependent selection [2], [3], fluctuating selection [2], [4], [5], selection maintained through segregation distortion [6], and selection maintaining more than two allelic classes [6]. Though these modes of balancing selection exhibit different evolutionary dynamics, they all lead to increased diversity around the site under selection, and therefore a decay in the density of polymorphisms with increasing genetic distance from the selected site. It is this information that
introduced in this article is that two allelic classes at a selected site are maintained for an infinitely long period of time at a constant equilibrium frequency by balancing selection. However, balancing selection is not restricted to act only on two stable allelic classes, and the equilibrium frequency can fluctuate with time and space. Examples of balancing selection that do not conform to our model assumptions are frequency-dependent selection [2], [3], fluctuating selection [2], [4], [5], selection maintained through segregation distortion [6], and selection maintaining more than two allelic classes [6]. Though these modes of balancing selection exhibit different evolutionary dynamics, they all lead to increased diversity around the site under selection, and therefore a decay in the density of polymorphisms with increasing genetic distance from the selected site. It is this information that 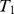 and
and 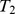 are employing to identify signatures of balancing selection, and though the dynamics of these modes of balancing selection violate the assumptions of our methods, it is likely that the statistics developed here could identify genomic signatures left behind by these selective scenarios provided selection was strong enough.
are employing to identify signatures of balancing selection, and though the dynamics of these modes of balancing selection violate the assumptions of our methods, it is likely that the statistics developed here could identify genomic signatures left behind by these selective scenarios provided selection was strong enough.
Within our scan, we identified a gene called FANK1, which is expressed during the transition from diploid to haploid states in meiosis [32], [33], is often identified as spermatogenesis-specific [32], [33], suppresses apoptosis [33], is imprinted [36], and exhibits evidence of segregation distortion (Fig. 8) [37]. These characteristics suggest that maintenance of polymorphism at FANK1 results from segregation distortion, which can occur when the allele favored by distortion is associated with negative fitness effects, particularly if the negative effect is pronounced in the homozygous state (see p. 562–563 of Charlesworth and Charlesworth [6]; Úbeda and Haig [54]). The distorting allele will increase in frequency when rare because of the segregation distortion in heterozygotes. But when it becomes common, selection will act against it because it will more often occur in the homozygous state when rare. Under such a scenario, theoretical results suggest that it is possible for a distorter to spread through a population without reaching fixation, obtaining a frequency that permits the maintenance of a stable polymorphism (see p. 564 of Charlesworth and Charlesworth [6]). In addition, the inclusion of imprinting at such a locus further enchances the parameter space at which a polymorphism can be maintained [54].
The function of FANK1 makes it a particularly good candidate for harboring alleles causing segregation distortion. It is expressed primarily during meiosis and inhibits apoptosis, which has previously been hypothesized to be associated with segregation distortion [55], [56]. A large proportion of sperm cells are eliminated by apoptosis, so allelic variants causing avoidance of apoptosis after meiosis could serve as segregation distorters. However, mutations that lead to avoidance of apoptosis may be associated with negative fitness effects, especially in the homozygous states, because they could lead to dysspermia or azoospermia. Apoptosis during spermatogenesis plays a critical role in maintaining the optimal relationship between the number of developing sperm cells and sertoli cells, which support developing sperm cells.
Though some of the sites identified in FANK1 show marginal levels of segregation distortion, the region displaying the largest level of segregation distortion in the human genome is located 300 kb upstream of FANK1 [37]. Further, a recent genome-wide association study for male fertility identified a significant SNP (rs9422913) located approximately 250 kb upstream of FANK1 [57]. Even though these regions are quite distant from FANK1, if strong enough linkage exists with FANK1, then it is possible for a two-locus segregation distorter to spread within a population (p. 569 of Charlesworth and Charlesworth [6]). Hence the signals of segregation distortion [37] and fertility [57] displayed in these regions upstream of FANK1 could be a result of an association with FANK1.
Thus, FANK1 is an interesting candidate for further study of balancing selection. The association of segregation distortion and balancing selection has been empirically described in other species, e.g., Caenorhabditis elegans [58]. However, as it has not yet been documented in humans, FANK1 may be the first example of a segregation distorter causing balancing selection in humans. However, further experiments would be needed to test the hypothesis of segregation distortion in FANK1.
In the last several years, there has been an accumulation of evidence against the pervasiveness of hard sweeps in some species, e.g., in humans [11]–[13]. Instead, other adaptive forces, such as balancing selection, could play an important role in shaping genetic variation across the genome. Interestingly, a recent theoretical study showed that a large proportion of adaptive mutations in diploids leads to heterozygote advantage [59], suggesting that much of the genome may be under balancing selection. If this intriguing prospect is true, then because our methods for detecting balancing selection are the most powerful that have been developed to date, they will be useful tools in uncovering the potentially many regions under balancing selection in humans and other species.
Materials and Methods
Estimating inter-species expected coalescence times
To compute the probabilities of polymorphism 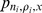 and substitution
and substitution 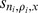 under our model, we must first obtain an estimate of the inter-species coalescent times
under our model, we must first obtain an estimate of the inter-species coalescent times  . For the purposes of our simulation and empirical analyses, we introduce a basic estimate (
. For the purposes of our simulation and empirical analyses, we introduce a basic estimate ( ) of the expected coalescence time between the ingroup and outgroup species. Consider a sample of
) of the expected coalescence time between the ingroup and outgroup species. Consider a sample of  lineages (i.e.,
lineages (i.e.,  haploid individuals) from an ingroup species and one lineage from an outgroup species. For simplicity, assume that the ingroup species, outgroup species, and ancestral species from which the ingroup and outgroup diverged has an effective population size of
haploid individuals) from an ingroup species and one lineage from an outgroup species. For simplicity, assume that the ingroup species, outgroup species, and ancestral species from which the ingroup and outgroup diverged has an effective population size of 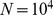 diploid individuals. Further, assume that the per-site per-generation mutation rate is
diploid individuals. Further, assume that the per-site per-generation mutation rate is 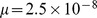 and that the total sequence length analyzed is
and that the total sequence length analyzed is  . We estimate the expected coalescence time of all
. We estimate the expected coalescence time of all  lineages in the ingroup species as
lineages in the ingroup species as 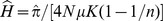 , where
, where 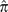 is the mean number of pairwise sequence differences and
is the mean number of pairwise sequence differences and 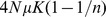 is the expected number of mutations for a sequence of length
is the expected number of mutations for a sequence of length 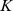 and
and  sampled lineages. Suppose that
sampled lineages. Suppose that 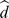 is the number of substitutions of fixed differences observed between the ingroup and outgroup species. Then we estimate the mean coalescence time between the ingroup and outgroup species by
is the number of substitutions of fixed differences observed between the ingroup and outgroup species. Then we estimate the mean coalescence time between the ingroup and outgroup species by 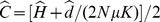 .
.
Application of the new test statistics to data
In the empirical analysis of human genomic data, we obtained values for the  and
and  test statistics for a large number of positions spaced across the genome. From these values, we overlapped protein coding regions (or genes including exons and introns) with the positions in the genome that the test statistics were calculated at. We assigned the value of the test statistic for the gene as the maximal value of the test statistic for the positions that it overlapped. We then ranked the set of genes based on their scores to identify genes that are outliers. Note that we are not attempting to identify regions with statistical significance or a certain
test statistics for a large number of positions spaced across the genome. From these values, we overlapped protein coding regions (or genes including exons and introns) with the positions in the genome that the test statistics were calculated at. We assigned the value of the test statistic for the gene as the maximal value of the test statistic for the positions that it overlapped. We then ranked the set of genes based on their scores to identify genes that are outliers. Note that we are not attempting to identify regions with statistical significance or a certain  -value threshold, but instead are looking for genes that may be outliers, and so the
-value threshold, but instead are looking for genes that may be outliers, and so the 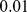 ,
, 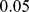 ,
, 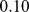 , and
, and 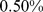 empirical cutoffs are not meant to represent a formal significance cutoff.
empirical cutoffs are not meant to represent a formal significance cutoff.
When applying the 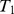 and
and 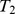 test statistics to simulated and empirical data, we do not estimate the rate of mutation
test statistics to simulated and empirical data, we do not estimate the rate of mutation 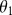 from
from 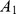 alleles to
alleles to 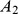 alleles or the rate of mutation
alleles or the rate of mutation 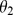 from
from 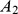 alleles to
alleles to 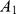 alleles at the selected site
alleles at the selected site 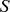 , as defined in the Hudson-Darden-Kaplan model. We instead treat these rates as a constant, with
, as defined in the Hudson-Darden-Kaplan model. We instead treat these rates as a constant, with 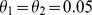 for the analyses in this article. The motivation is that, if these mutation rates did not exist, then the tree height would increase rapidly for small recombination rates. Our method assumes that a most recent common ancestor of the set of sampled alleles is reached more recently than the inter-species coalescence time
for the analyses in this article. The motivation is that, if these mutation rates did not exist, then the tree height would increase rapidly for small recombination rates. Our method assumes that a most recent common ancestor of the set of sampled alleles is reached more recently than the inter-species coalescence time 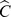 between the ingroup and outgroup species (i.e.,
between the ingroup and outgroup species (i.e., 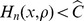 even for small
even for small 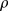 ). Simulation results (see Evaluating the methods using simulations) show that our new methods perform extremely well, even though we set the nuisance
). Simulation results (see Evaluating the methods using simulations) show that our new methods perform extremely well, even though we set the nuisance 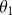 and
and 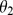 parameters to a constant value. To maximize of the equilibrium frequency
parameters to a constant value. To maximize of the equilibrium frequency  of the
of the 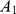 allele, we utilized the value of
allele, we utilized the value of  , denoted by
, denoted by 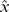 , that maximized the composite likelihood under the model, by choosing
, that maximized the composite likelihood under the model, by choosing 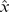 from values of
from values of 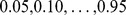 .
.
Simulation procedure to evaluate the performance of  and
and 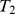
We applied 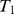 and
and 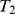 to data simulated under population divergence models, using parameters to mimic humans (ingroup) and chimpanzees (outgroup). The models that we simulated under are illustrated in Figure 2. For each of three models, we set each of the ingroup, outgroup, and ancestral population sizes to
to data simulated under population divergence models, using parameters to mimic humans (ingroup) and chimpanzees (outgroup). The models that we simulated under are illustrated in Figure 2. For each of three models, we set each of the ingroup, outgroup, and ancestral population sizes to 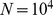 diploid individuals [60] and the divergence time between the ingroup and the outgroup species to
diploid individuals [60] and the divergence time between the ingroup and the outgroup species to 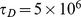 years ago [61]. We assumed a generation time of 20 years [62], a mutation rate of
years ago [61]. We assumed a generation time of 20 years [62], a mutation rate of 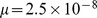 mutations per-nucleotide per-generation [62], a recombination rate of
mutations per-nucleotide per-generation [62], a recombination rate of 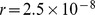 recombinations per-nucleotide per-generation, and a sequence length of
recombinations per-nucleotide per-generation, and a sequence length of 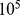 nucleotides. Assuming a per-generation selection coefficient
nucleotides. Assuming a per-generation selection coefficient  , where
, where 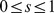 , and a dominance parameter
, and a dominance parameter 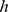 , where
, where 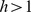 , at time
, at time 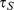 , a selected allele arose and evolved under an overdominance model with
, a selected allele arose and evolved under an overdominance model with 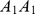 homozygotes having fitness 1,
homozygotes having fitness 1, 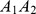 heterozygotes having fitness
heterozygotes having fitness 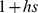 , and
, and 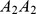 homozygotes having fitness
homozygotes having fitness  . The formulation of this overdominance model is similar to that of [63] in which the fitness is
. The formulation of this overdominance model is similar to that of [63] in which the fitness is 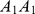 is 1,
is 1, 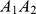 is
is 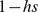 , and
, and 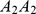 is
is  . Under the Gillespie formulation, overdominance occurs when
. Under the Gillespie formulation, overdominance occurs when 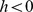 , whereas it occurs when
, whereas it occurs when 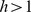 in our formulation. However, both result in an equilibrium frequency of
in our formulation. However, both result in an equilibrium frequency of 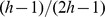 . Simulations were performed using mpop
[64], which was seeded with population-level chromosome data generated by the neutral coalescent simulator ms
[65]. After the completion of each simulation, we sampled 18 chromosomes from the ingroup species and one chromosome from the outgroup species. For each set of parameter values, we simulated
. Simulations were performed using mpop
[64], which was seeded with population-level chromosome data generated by the neutral coalescent simulator ms
[65]. After the completion of each simulation, we sampled 18 chromosomes from the ingroup species and one chromosome from the outgroup species. For each set of parameter values, we simulated 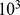 independent replicates. Ancestral alleles were called using the outgroup species, and so the called ancestral allele may not actually be the true ancestral allele. For each of the three demographic scenarios, we set
independent replicates. Ancestral alleles were called using the outgroup species, and so the called ancestral allele may not actually be the true ancestral allele. For each of the three demographic scenarios, we set 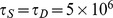 years ago. For the bottleneck model (Fig. 2B
), we set the bottleneck population size to
years ago. For the bottleneck model (Fig. 2B
), we set the bottleneck population size to 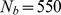 diploid individuals, the time at which the bottleneck began to
diploid individuals, the time at which the bottleneck began to 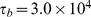 years ago, and the time at which the bottleneck ended to
years ago, and the time at which the bottleneck ended to 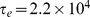 years ago [66], [67]. For the growth model (Fig. 2C
), we set the expanded population size to
years ago [66], [67]. For the growth model (Fig. 2C
), we set the expanded population size to 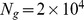 diploid individuals and the time at which the population began to grow to
diploid individuals and the time at which the population began to grow to 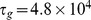 years ago [67]. Additionally, we considered a more ancient balanced polymorphism arising
years ago [67]. Additionally, we considered a more ancient balanced polymorphism arising 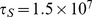 years ago and a more recent balanced polymorphism arising
years ago and a more recent balanced polymorphism arising 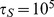 years ago. Because the forward simulations in mpop are computationally burdensome, we rescaled appropriate parameters by a factor of 10 such that the scaled population parameters remain the same, but the simulations are substantially sped up (by approximately a factor of
years ago. Because the forward simulations in mpop are computationally burdensome, we rescaled appropriate parameters by a factor of 10 such that the scaled population parameters remain the same, but the simulations are substantially sped up (by approximately a factor of 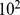 ). Note that scaling parameters in this way can somewhat affect the time to fixation of selected alleles. The distribution of false positive rates was generated by
). Note that scaling parameters in this way can somewhat affect the time to fixation of selected alleles. The distribution of false positive rates was generated by 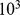 replicate neutral simulations from mpop, using the same parameters as the corresponding selection scenarios (including the rescaling factor) except without introducing a selected allele.
replicate neutral simulations from mpop, using the same parameters as the corresponding selection scenarios (including the rescaling factor) except without introducing a selected allele.
Matching the density of polymorphic sites
In the current set of simulations, the bottleneck and growth models each produce a different density of polymorphisms (i.e., number of segregating sites) than the constant size model. This section seeks to find an ancestral effective size for the growth and the bottleneck models such that the mean density of polymorphisms is close to that of the constant size model. We use eq. 1 in Marth et al. (2004) [68] to calculate the expected frequency spectrum under the bottleneck and growth models. The equation is
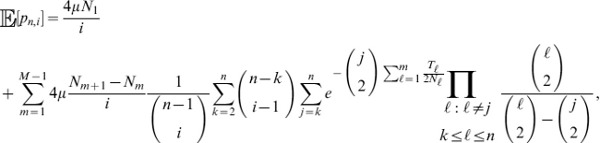 |
(15) |
where 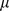 is the per-generation mutation rate,
is the per-generation mutation rate, 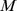 is the number of epochs,
is the number of epochs, 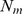 for
for 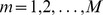 , is the effective population size for epoch
, is the effective population size for epoch  , and
, and 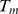 for
for 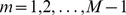 , is the duration of time spent in epoch
, is the duration of time spent in epoch  . Our growth model contains two epochs, and so the appropriate version of the equation is when
. Our growth model contains two epochs, and so the appropriate version of the equation is when  . Setting the number of epochs to two, we the expected frequency spectrum under the growth model as
. Setting the number of epochs to two, we the expected frequency spectrum under the growth model as
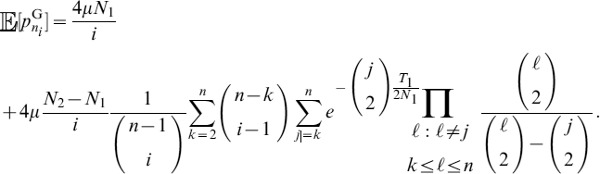 |
(16) |
Note that in our growth model, 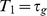 ,
, 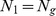 , and
, and 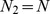 . Denote the ratio of effective size during growth to the ancestral effective size as
. Denote the ratio of effective size during growth to the ancestral effective size as 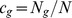 . Then we can rewrite the equation as
. Then we can rewrite the equation as
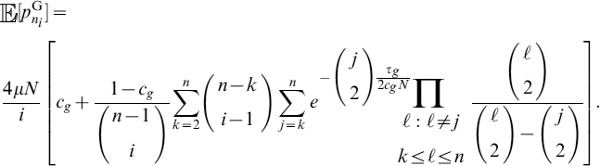 |
(17) |
Consider an ancestral reference effective size  (
(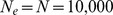 for the constant size model). Denote the expected number of segregating sites in a constant size model, conditional on effective size
for the constant size model). Denote the expected number of segregating sites in a constant size model, conditional on effective size 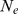 as
as 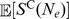 . Conditional on this ancestral reference effective size
. Conditional on this ancestral reference effective size 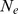 , the expected site frequency spectrum under our growth model is
, the expected site frequency spectrum under our growth model is
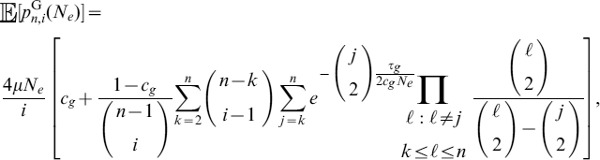 |
(18) |
where  under our growth model. Therefore, the expected number of segregating sites conditional on reference effective size
under our growth model. Therefore, the expected number of segregating sites conditional on reference effective size  is
is  . We obtain a growth model that produces the same density of polymorphic sites as our constant size model by choosing
. We obtain a growth model that produces the same density of polymorphic sites as our constant size model by choosing
| (19) |
Our bottleneck model contains three epochs, and so the appropriate version of the equation is when 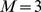 . Setting the number of epochs to three, we the expected frequency spectrum under the bottleneck model as
. Setting the number of epochs to three, we the expected frequency spectrum under the bottleneck model as
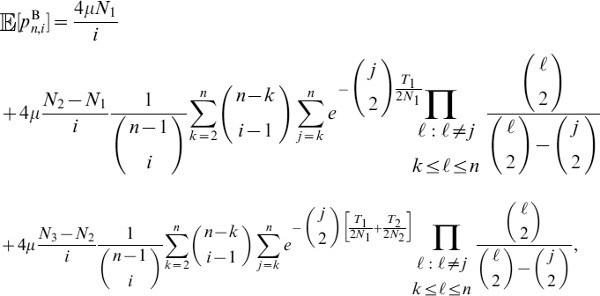 |
(20) |
Note that in our bottleneck model, 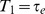 ,
, 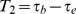 ,
, 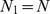 ,
, 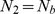 , and
, and 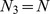 . Denote the ratio of the effective size during the bottleneck to the ancestral effective size as
. Denote the ratio of the effective size during the bottleneck to the ancestral effective size as 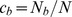 . Then we can rewrite the equation as
. Then we can rewrite the equation as
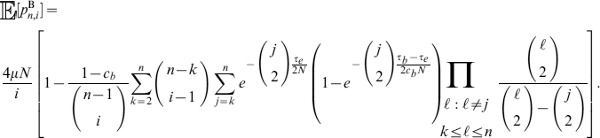 |
(21) |
Conditional on this reference effective size, the expected site frequency spectrum under our bottleneck model is
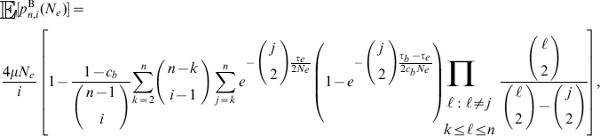 |
(22) |
where  under our bottleneck model. Therefore, the expected number of segregating sites conditional on reference effective size
under our bottleneck model. Therefore, the expected number of segregating sites conditional on reference effective size  is
is  . We obtain a bottleneck model that produces the same density of polymorphic sites as our constant size model by choosing
. We obtain a bottleneck model that produces the same density of polymorphic sites as our constant size model by choosing
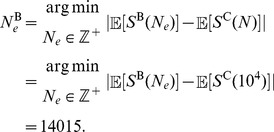 |
(23) |
Empirical dataset construction
We used data from nine European and nine African diploid genomes sequenced by Complete Genomics [69]. All individuals were unrelated [70], with the European individuals from the CEU population (NA06985, NA06994, NA07357, NA10851, NA12004, NA12889, NA12890, NA12891, NA12892) and the African individuals from the YRI population (NA18501, NA18502, NA18504, NA18505, NA18508, NA18517, NA19129, NA19238, NA19329). We used the genotype calls made by Complete Genomics that were found in the “masterVarBeta” files. We downloaded pairwise alignments between human reference hg18 and chimpanzee reference panTro2 from the UCSC Genome Browser at http://genome.ucsc.edu/. Sites with more than two distinct alleles across all Complete Genomics individuals as well as the hg18-panTro2 alignments, sites in the Complete Genomics data where one of the two alleles did not match the reference sequence, and sites that were within two nucleotides of structural variants called in any one of the Complete Genomics individuals were removed. In addition, combining all 54 unrelated individuals in the Complete Genomics dataset, sites that had a  -value less than
-value less than 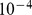 for a one-tailed Hardy-Weinberg test of excess heterozygotes [71] were excluded. We used the full set of 54 unrelated individuals, totalling 108 alleles, so that we would have sufficient power to detect Hardy-Weinberg departures due to excess heterozygotes. Sites flagged as departing from Hardy-Weinberg proportions in this set of 54 individuals were then filtered out in the smaller subsets of nine CEU and nine YRI individuals. It should be noted that under a scenario of heterozygote advantage, it is expected that we should observe an excess of heterozygous individuals at sites in the vicinity of the site under balancing selection. However, a major concern with sequencing data are mapping errors, and so the Hardy-Weinberg filter is necessary to reduce the confounding effects of regions with these bioinformatical artifacts. As a consequence, this filter may increase the chance that we miss certain regions that are under balancing selection in our scan. Finally, sites that were polymorphic in the Complete Genomics sample (i.e., either CEU or YRI) and sites that contained a fixed difference between the Complete Genomics sample and the chimpanzee reference sequence were retained. As in the simulations, the ancestral allele was called using the chimpanzee outgroup, and so the called ancestral allele may not be the true ancestral allele. However, simulation results shows that our new methods perform well even when the ancestral allele is potentially misspecified. Further, it may be possible to account for ancestral allele misspecification by using multiple outgroups, or by statistically accounting for the misspecification [72].
for a one-tailed Hardy-Weinberg test of excess heterozygotes [71] were excluded. We used the full set of 54 unrelated individuals, totalling 108 alleles, so that we would have sufficient power to detect Hardy-Weinberg departures due to excess heterozygotes. Sites flagged as departing from Hardy-Weinberg proportions in this set of 54 individuals were then filtered out in the smaller subsets of nine CEU and nine YRI individuals. It should be noted that under a scenario of heterozygote advantage, it is expected that we should observe an excess of heterozygous individuals at sites in the vicinity of the site under balancing selection. However, a major concern with sequencing data are mapping errors, and so the Hardy-Weinberg filter is necessary to reduce the confounding effects of regions with these bioinformatical artifacts. As a consequence, this filter may increase the chance that we miss certain regions that are under balancing selection in our scan. Finally, sites that were polymorphic in the Complete Genomics sample (i.e., either CEU or YRI) and sites that contained a fixed difference between the Complete Genomics sample and the chimpanzee reference sequence were retained. As in the simulations, the ancestral allele was called using the chimpanzee outgroup, and so the called ancestral allele may not be the true ancestral allele. However, simulation results shows that our new methods perform well even when the ancestral allele is potentially misspecified. Further, it may be possible to account for ancestral allele misspecification by using multiple outgroups, or by statistically accounting for the misspecification [72].
To obtain recombination rates between pairs of sites, we used the sex-averaged pedigree-based human recombination map from deCODE Genetics [73]. We constructed recombination rates between all pairs of sites in the filtered Complete Genomics samples by linearly interpolating rates between adjacent sites within the sex-averaged maps.
Supporting Information
Performance of  ,
,  , HKA, and Tajima's
, HKA, and Tajima's 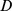 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 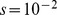 and dominance parameter
and dominance parameter  . Each row represents a different
. Each row represents a different  value. The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
value. The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Mean difference in the number of polymorphic sites for a model with 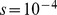 versus one with
versus one with 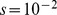 as a function of the distance from the site under balancing selection. Simulations were performed under the constant size divergence model in Figure 2A
with selection parameter
as a function of the distance from the site under balancing selection. Simulations were performed under the constant size divergence model in Figure 2A
with selection parameter  , dominance parameter
, dominance parameter 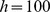 , and time of selection
, and time of selection 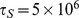 years ago. The mean difference in polymorphic sites is calculated for bins of size one kilobase and is plotted for 50 bins.
years ago. The mean difference in polymorphic sites is calculated for bins of size one kilobase and is plotted for 50 bins.
(PDF)
Performance of 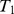 ,
, 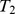 , HKA, and Tajima's
, HKA, and Tajima's 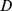 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 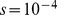 and dominance parameter
and dominance parameter 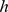 . Each row represents a different
. Each row represents a different 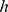 value. The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
value. The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of 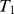 ,
, 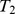 , HKA, and Tajima's
, HKA, and Tajima's 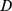 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 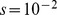 and dominance parameter
and dominance parameter 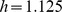 . The first panel is the divergence model in Figure 2A
. The second panel is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third panel is the divergence model in Figure 2C
with recent population growth within the ingroup species.
. The first panel is the divergence model in Figure 2A
. The second panel is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third panel is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of 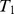 and
and 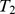 under the constant size divergence model in Figure 2A
with no selected allele (neutrality). The first and second panels are scenarios in which we have erroneously over-estimated the recombination rate by two and one orders of magnitude, respectively (i.e., we respectively assumed recombination rates of
under the constant size divergence model in Figure 2A
with no selected allele (neutrality). The first and second panels are scenarios in which we have erroneously over-estimated the recombination rate by two and one orders of magnitude, respectively (i.e., we respectively assumed recombination rates of 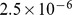 and
and 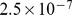 per base per generation when the simulations were performed using a rate of
per base per generation when the simulations were performed using a rate of 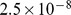 per base per generation). The third and fourth panels are scenarios in which we have erroneously under-estimated the recombination rate by one and two orders of magnitude, respectively (i.e., we respectively assumed recombination rates of
per base per generation). The third and fourth panels are scenarios in which we have erroneously under-estimated the recombination rate by one and two orders of magnitude, respectively (i.e., we respectively assumed recombination rates of 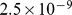 and
and 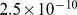 per base per generation when the simulations were performed using a rate of
per base per generation when the simulations were performed using a rate of 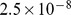 per base per generation). False positive rate is determined by neutral simulations under a model with recombination rate of
per base per generation). False positive rate is determined by neutral simulations under a model with recombination rate of 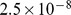 per base per generation.
per base per generation.
(PDF)
Performance of 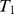 and
and 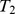 under the constant size divergence model in Figure 2A
with selection parameter
under the constant size divergence model in Figure 2A
with selection parameter 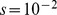 , dominance parameter
, dominance parameter 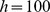 or 1.5, and time of selection
or 1.5, and time of selection 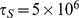 years ago. The first and second columns are scenarios in which we have erroneously over-estimated the recombination rate by two and one orders of magnitude, respectively (i.e., we respectively assumed recombination rates of
years ago. The first and second columns are scenarios in which we have erroneously over-estimated the recombination rate by two and one orders of magnitude, respectively (i.e., we respectively assumed recombination rates of 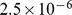 and
and 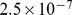 per base per generation when the simulations were performed using a rate of
per base per generation when the simulations were performed using a rate of 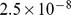 per base per generation). The third and fourth columns are scenarios in which we have erroneously under-estimated the recombination rate by one and two orders of magnitude, respectively (i.e., we respectively assumed recombination rates of
per base per generation). The third and fourth columns are scenarios in which we have erroneously under-estimated the recombination rate by one and two orders of magnitude, respectively (i.e., we respectively assumed recombination rates of 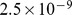 and
and 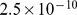 per base per generation when the simulations were performed using a rate of
per base per generation when the simulations were performed using a rate of 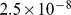 per base per generation). False positive rate is determined by neutral simulations under a model with recombination rate of
per base per generation). False positive rate is determined by neutral simulations under a model with recombination rate of 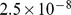 per base per generation.
per base per generation.
(PDF)
Demographic models used in simulations in which a selected allele arises prior to the split a pair of species. (A) Divergence model. Model parameters are a diploid effective population size 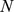 , divergence time
, divergence time 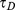 of the ingroup and outgroup species, and the time
of the ingroup and outgroup species, and the time 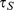 when the selected allele arises. (B) Divergence model with a recent bottleneck within the ingroup species. Additional model parameters are the diploid effective population size
when the selected allele arises. (B) Divergence model with a recent bottleneck within the ingroup species. Additional model parameters are the diploid effective population size 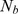 during the bottleneck, the time
during the bottleneck, the time 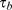 when the bottleneck began, and the time
when the bottleneck began, and the time 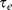 when the bottleneck ended. (C) Divergence model with recent population growth within the ingroup species. Additional model parameters are the current diploid effective population size
when the bottleneck ended. (C) Divergence model with recent population growth within the ingroup species. Additional model parameters are the current diploid effective population size 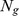 after recent growth and the time
after recent growth and the time 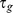 when the growth occurred.
when the growth occurred.
(PDF)
Performance of 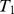 ,
, 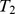 , HKA, and Tajima's
, HKA, and Tajima's 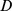 under the demographic models in Figure S7 with selection parameter
under the demographic models in Figure S7 with selection parameter 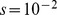 and dominance parameter
and dominance parameter 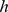 . Each row represents a different
. Each row represents a different 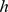 value. The first column is the divergence model in Figure S7
A. The second column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
value. The first column is the divergence model in Figure S7
A. The second column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
(PDF)
Performance of 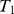 ,
, 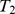 , HKA, and Tajima's
, HKA, and Tajima's 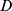 under the demographic models in Figure S7 with selection parameter
under the demographic models in Figure S7 with selection parameter 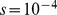 and dominance parameter
and dominance parameter  . Each row represents a different
. Each row represents a different  value. The first column is the divergence model in Figure S7
A. The second column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
value. The first column is the divergence model in Figure S7
A. The second column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
(PDF)
Performance of 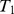 ,
, 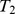 , HKA, and Tajima's
, HKA, and Tajima's 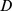 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 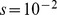 , dominance parameter
, dominance parameter 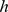 , and time of selection
, and time of selection 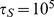 . The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
. The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of 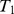 ,
, 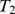 , HKA, and Tajima's
, HKA, and Tajima's 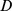 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 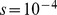 , dominance parameter
, dominance parameter 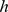 , and time of selection
, and time of selection 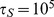 . The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
. The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of 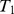 ,
, 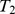 , HKA, and Tajima's
, HKA, and Tajima's 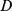 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 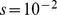 and dominance parameter
and dominance parameter 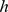 . Each row represents a different
. Each row represents a different 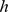 value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size
value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size 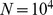 individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of 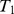 ,
, 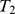 , HKA, and Tajima's
, HKA, and Tajima's 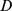 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 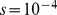 and dominance parameter
and dominance parameter 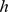 . Each row represents a different
. Each row represents a different 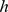 value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size
value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size 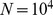 individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of 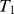 ,
, 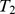 , HKA, and Tajima's
, HKA, and Tajima's 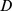 under the demographic models in Figure S7 with selection parameter
under the demographic models in Figure S7 with selection parameter 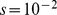 and dominance parameter
and dominance parameter  . Each row represents a different
. Each row represents a different  value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size
value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size 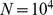 individuals. The first column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
individuals. The first column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
(PDF)
Performance of 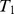 ,
, 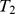 , HKA, and Tajima's
, HKA, and Tajima's 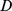 under the demographic models in Figure S7 with selection parameter
under the demographic models in Figure S7 with selection parameter 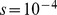 and dominance parameter
and dominance parameter 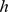 . Each row represents a different
. Each row represents a different 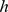 value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size
value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size 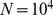 individuals. The first column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
individuals. The first column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
(PDF)
Performance of 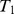 ,
, 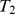 , HKA, and Tajima's
, HKA, and Tajima's 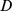 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 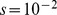 , and dominance parameter
, and dominance parameter 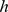 , and time of selection
, and time of selection 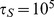 . Each row represents a different
. Each row represents a different 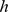 value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size
value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size 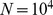 individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of 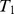 ,
, 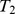 , HKA, and Tajima's
, HKA, and Tajima's 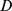 under the demographic models in Figure 2 with selection parameter
under the demographic models in Figure 2 with selection parameter 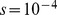 , and dominance parameter
, and dominance parameter 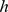 , and time of selection
, and time of selection 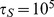 . Each row represents a different
. Each row represents a different 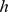 value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size
value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size 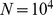 individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Manhattan plot of genome-wide scans for balancing selection within the CEU population using the 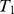 test statistic. From bottom to top, the horizontal dotted gray lines indicate the
test statistic. From bottom to top, the horizontal dotted gray lines indicate the 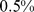 ,
, 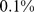 ,
, 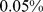 , and
, and 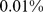 empirical cutoffs, respectively. The
empirical cutoffs, respectively. The 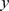 -axis is truncated at log composite likelihood ratio of zero.
-axis is truncated at log composite likelihood ratio of zero.
(PDF)
Manhattan plot of genome-wide scans for balancing selection within the YRI population using the 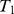 test statistic. From bottom to top, the horizontal dotted gray lines indicate the
test statistic. From bottom to top, the horizontal dotted gray lines indicate the 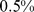 ,
, 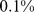 ,
, 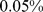 , and
, and 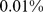 empirical cutoffs, respectively. The
empirical cutoffs, respectively. The 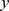 -axis is truncated at log composite likelihood ratio of zero.
-axis is truncated at log composite likelihood ratio of zero.
(PDF)
Manhattan plot of genome-wide scans for balancing selection within the CEU population using the 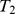 test statistic. From bottom to top, the horizontal dotted gray lines indicate the
test statistic. From bottom to top, the horizontal dotted gray lines indicate the 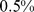 ,
, 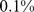 ,
, 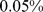 , and
, and 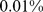 empirical cutoffs, respectively. The
empirical cutoffs, respectively. The 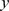 -axis is truncated at log composite likelihood ratio of zero.
-axis is truncated at log composite likelihood ratio of zero.
(PDF)
Manhattan plot of genome-wide scans for balancing selection within the YRI population using the 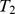 test statistic. From bottom to top, the horizontal dotted gray lines indicate the
test statistic. From bottom to top, the horizontal dotted gray lines indicate the 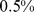 ,
, 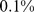 ,
, 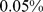 , and
, and 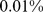 empirical cutoffs, respectively. The
empirical cutoffs, respectively. The 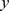 -axis is truncated at log composite likelihood ratio of zero.
-axis is truncated at log composite likelihood ratio of zero.
(PDF)
Signals of balancing selection within the HLA region for the CEU (blue) and YRI (orange) populations using the 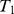 test statistic. From bottom to top, the horizontal dotted gray lines indicate the
test statistic. From bottom to top, the horizontal dotted gray lines indicate the  ,
, 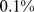 ,
, 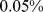 , and
, and 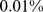 empirical cutoffs, respectively.
empirical cutoffs, respectively.
(PDF)
Signal of balancing selection at the FANK1 gene for the CEU (blue) and YRI (orange) populations using the 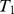 test statistic. From bottom to top, the horizontal dotted gray lines indicate the
test statistic. From bottom to top, the horizontal dotted gray lines indicate the 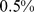 ,
, 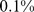 ,
, 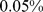 , and
, and 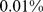 empirical cutoffs, respectively. SNPs (rsIDs) correspond to markers showing significant levels of transmission distortion within the Meyer et al. study.
empirical cutoffs, respectively. SNPs (rsIDs) correspond to markers showing significant levels of transmission distortion within the Meyer et al. study.
(PDF)
Signal of balancing selection at the FANK1 gene for the CEU (blue) and YRI (orange) populations when removing either 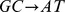 transitions or all transitions. SNPs (rsIDs) correspond to markers showing significant levels of transmission distortion within the Meyer et al. study.
transitions or all transitions. SNPs (rsIDs) correspond to markers showing significant levels of transmission distortion within the Meyer et al. study.
(PDF)
Genealogy at the site of balancing selection.
(PDF)
Haplotype trees based on randomly sampling 18 haplotypes without replacement from a random simulation under the model in Figure S7
A. Trees were generated using UPGMA applied to a distance matrix of the proportion of nucleotide differences between each pair of haplotypes. The  -kilobase (kb) window represents a region that is
-kilobase (kb) window represents a region that is  kb in length and is centered in the middle of the haplotype.
kb in length and is centered in the middle of the haplotype.
(PDF)
Top 100 signals in the CEU population using the 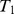 test statistic.
test statistic.
(PDF)
Top 100 signals in the YRI population using the 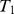 test statistic.
test statistic.
(PDF)
Top 100 signals in the CEU population using the 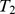 test statistic.
test statistic.
(PDF)
Top 100 signals in the YRI population using the 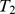 test statistic.
test statistic.
(PDF)
GO process analysis of top 100 signals, when compared to all signals, from CEU population using the 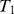 test statistic.
test statistic.
(PDF)
GO process analysis of top 100 signals, when compared to all signals, from YRI population using the 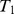 test statistic.
test statistic.
(PDF)
GO process analysis of top 100 signals, when compared to all signals, from CEU population using the 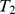 test statistic.
test statistic.
(PDF)
GO process analysis of top 100 signals, when compared to all signals, from YRI population using the 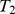 test statistic.
test statistic.
(PDF)
GO function analysis of top 100 signals, when compared to all signals, from CEU population using the 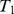 test statistic.
test statistic.
(PDF)
GO function analysis of top 100 signals, when compared to all signals, from YRI population using the 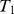 test statistic.
test statistic.
(PDF)
GO function analysis of top 100 signals, when compared to all signals, from CEU population using the 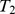 test statistic.
test statistic.
(PDF)
GO component analysis of top 100 signals, when compared to all signals, from CEU population using the 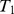 test statistic.
test statistic.
(PDF)
GO component analysis of top 100 signals, when compared to all signals, from YRI population using the 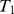 test statistic.
test statistic.
(PDF)
GO component analysis of top 100 signals, when compared to all signals, from CEU population using the 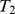 test statistic.
test statistic.
(PDF)
GO component analysis of top 100 signals, when compared to all signals, from YRI population using the 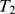 test statistic.
test statistic.
(PDF)
GO process analysis of ranked signals from CEU population using the 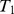 test statistic.
test statistic.
(PDF)
GO process analysis of ranked signals from YRI population using the 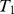 test statistic.
test statistic.
(PDF)
GO process analysis of ranked signals from CEU population using the 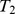 test statistic.
test statistic.
(PDF)
GO process analysis of ranked signals from YRI population using the 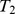 test statistic.
test statistic.
(PDF)
GO function analysis of ranked signals from CEU population using the 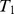 test statistic.
test statistic.
(PDF)
GO function analysis of ranked signals from YRI population using the 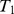 test statistic.
test statistic.
(PDF)
GO function analysis of ranked signals from CEU population using the 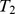 test statistic.
test statistic.
(PDF)
GO function analysis of ranked signals from YRI population using the 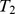 test statistic.
test statistic.
(PDF)
GO component analysis of ranked signals from CEU population using the 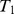 test statistic.
test statistic.
(PDF)
GO component analysis of ranked signals from YRI population using the 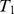 test statistic.
test statistic.
(PDF)
GO component analysis of ranked signals from CEU population using the 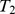 test statistic.
test statistic.
(PDF)
GO component analysis of ranked signals from YRI population using the 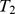 test statistic.
test statistic.
(PDF)
Acknowledgments
We thank four anonymous reviewers for their insightful comments, which significantly improved our manuscript. We also thank Zachary Szpiech for coming up with the name BALLET and Zelia Ferreira for help testing early versions of BALLET.
Funding Statement
This material was supported by National Science Foundation grant DBI-1103639 (MD), a Miller Research Fellowship from the Miller Research Institute at the University of California, Berkeley (KEL), and National Institutes of Health grant 3R01HG03229-07 (RN). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Fisher RA (1922) On the dominance ratio. Proc Roy Soc Edin 42: 321–341. [Google Scholar]
- 2.Andrés AM (2011) Balancing selection in the human genome. In: Encyclopedia of Life Sciences, Chichester: John Wiley and Sons. [Google Scholar]
- 3. Wilson DS, Turelli M (1986) Stable underdominance and the evolutionary invasion of empty niches. Am Nat 127: 835–850. [Google Scholar]
- 4. Levene H (1953) Genetic equilibrium when more than one ecological niche is available. Am Nat 83: 331–333. [Google Scholar]
- 5. Nagylaki T (1975) Polymorphisms in cyclically varying environments. Heredity 35: 67–74. [DOI] [PubMed] [Google Scholar]
- 6.Charlesworth B, Charlesworth D (2010) Elements of evolutionary genetics. Greenwood Village, CO: Roberts and Company Publishers. [Google Scholar]
- 7. Ségurel L, Thompson EE, Flutre T, Lovstad J, Venkat A, et al. (2012) The ABO blood group is a trans-species polymorphism in primates. Proc Natl Acad Sci USA 109: 18493–18498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Klein J, Satta Y, O'hUigín C (1993) The molecular descent of the major histocompatibility complex. Annu Rev Immunol 11: 269–95. [DOI] [PubMed] [Google Scholar]
- 9. Klein J, Sato A, Nagl S, O'hUigín C (1998) Molecular trans-species polymorphism. Annu Rev Ecol Syst 29: 1–21. [Google Scholar]
- 10. Klein J, Sato A, Nikolaidis N (2007) MHC, TSP, and the origin of species: from immunogenetics to evlutionary genetics. Annu Rev Genet 41: 281–304. [DOI] [PubMed] [Google Scholar]
- 11. Hernandez RD, Kelley JL, Elyashiv E, Melton SC, Auton A, et al. (2011) Classic selective sweeps were rare in recent human evolution. Science 331: 920–924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Lohmueller KE, Albrechtsen A, Li Y, Y KS, Korneliussen T, et al. (2011) Natural selection affects multiple aspects of genetic variation at putatively neutral sites across the human genome. PLoS Genet 7: e1002326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Granka JM, Henn BM, Gignoux CR, Kidd JM, Bustamante CD, et al. (2012) Limited evidence for classic selective sweeps in African populations. Genetics 92: 1049–64 doi:10.1534/genetics.112.144071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Bubb KL, Bovee D, Buckley D, Haugen E, Kibukawa M, et al. (2006) Scan of human genome reveals no new loci under ancient balancing selection. Genetics 173: 2165–2177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Andrés AM, Hubisz MJ, Indap A, Torgerson DG, Degenhardt JD, et al. (2009) Targets of balancing selection in the human genome. Mol Biol Evol 26: 2755–2764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Hudson RR, Kreitman M, Aguadé M (1987) A test of neutral marker evolution based on nucleotide data. Genetics 116: 153–159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Tajima F (1989) Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123: 585–595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Innan H (2006) Modified Hudson-Kreitman-Aguadé test and two-dimensional evaluation of neutrality tests. Genetics 173: 1725–1733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Leffler EM, Gao Z, Pfeifer S, Ségurel L, Auton A, et al. (2013) Multiple instances of ancient balancing selection shared between humans and chimpanzees. Science 339: 1578–1582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Kaplan NL, Darden T, Hudson RR (1988) The coalescent proces in models with selection. Genetics 120: 819–829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Hudson RR, Kaplan NL (1988) The coalescent process in models with selection and recombination. Genetics 120: 831–840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Hudson RR (2001) Two-locus sampling distributions and their application. Genetics 159: 1805–1817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Kim Y, Stephan W (2002) Detecting a local signature of genetic hitchhiking along a recombining chromosome. Genetics 160: 765–777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Kim Y, Nielsen R (2004) Linkage disequilibrium as a signature of selective sweeps. Genetics 167: 1513–1524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Jensen JD, Kim Y, DuMont VB, Aquadro CF, Bustamante CD (2005) Distinguishing between selective sweeps and demography using DNA polymorphism data. Genetics 170: 1401–1410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Nielsen R, Williamson S, Kim Y, Hubisz MJ, Clark AG, et al. (2005) Genomic scans for selective sweeps using SNP data. Genome Res 15: 1566–1575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Nielsen R, Hubsz MJ, Hellmann I, Torgerson D, Andrés AM, et al. (2009) Darwinian and demographic forces affecting human protein coding genes. Genome Res 19: 838–849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Chen H, Patterson N, Reich D (2010) Population differentiation as a test for selective sweeps. Genome Res 20: 393–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Thomas LH (1949) Elliptic problems in linear difference equations over a network. New York: Watson Sci. Comput. Lab. Rept., Columbia University. [Google Scholar]
- 30. Takahata N, Nei M (1990) Allelic genealogy under overdominant and frequency-dependent selection and polymorphism of major histocompatibility loci. Genetics 124: 967–978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Hedrick PW (2002) Pathogen resistance and geneic variation at MHC loci. Evolution 56: 1902–1908. [DOI] [PubMed] [Google Scholar]
- 32. Zheng Z, Zheng H, Yan W (2007) Fank1 is a testis-specific gene encoding a nuclear protein exclusively expressed during the transition from meiotic to the haploid phase of spermatogenesis. Gene Expr Patterns 7: 777–783. [DOI] [PubMed] [Google Scholar]
- 33. Wang H, Song W, Hu T, Zhang N, Miao S, et al. (2011) Fank1 interacts with Jab1 and regulates cell apoptosis via the AP-1 pathway. Cell Mol Life Sci 68: 2129–2139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Hwang KC, Park SY, Park SP, Lim JH, Cui XS, et al. (2005) Specific maternal transcripts in bovie oocytes and cleavaged embryos: identification with novel DDRT-PCR methods. Mol Reprod Dev 71: 275–283. [DOI] [PubMed] [Google Scholar]
- 35. Zuccotti M, Merico V, Sacchi L, Bellone M (2008) Brink R T C nd Bellazzi, (2008) et al. Maternal Oct-4 is a potential key regulator of the developmental compentence of mouse oocytes. BMC Dev Biol 8: 97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Li Y, Zhu J, Tian G, Li N, Li Q, et al. (2010) The DNA methylome of human peripheral blood mononuclear cells. PLoS Biol 8: e1000533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Meyer WK, Arbeithuber B, Ober C, Ebner T, Tiemann-Boege I, et al. (2012) Evaluating the evidence for transmission distortion in human pedigress. Genetics 191: 215–232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Akey JM, Eberle MA, Rieder MJ, Carlson CS, Shriver MD, et al. (2004) Population history and natural selection shape patterns of genetic variation in 132 genes. PLoS Biol 2: e286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Eden E, Lipson D, Yogev S, Yakhini Z (2007) Discovering motifs in ranked lists of DNA sequences. PLoS Comput Biol 3: e39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Eden E, Navon R, Steinfeld I, Lipson D, Yakhini Z (2009) GOrilla: a tool for discovery and visualization of enriched GO terms in ranked gene lists. BMC Bioinformatics 10: 48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Barton NH, Etheridge AM (2004) The effect of selection on genealogies. Genetics 166: 1115–1131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Barton NH, Etheridge AM, Sturm AK (2004) Coalescence in a random background. Ann Appl Probab 14: 754–785. [Google Scholar]
- 43. Green RE, Krause J, Briggs AW, Marici T, Stenzel U, et al. (2010) A draft sequence of the Neandertal genome. Science 328: 710–722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Reich D, Green RE, Kircher M, Krause J, Patterson N, et al. (2010) Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 468: 1053–1060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Jensen JD, Thornton KR, Bustamante CD, Aquadro CF (2007) On the utility of linkage disequilibrium as a statistic for identifying targets of positive selection in nonequilibrium populations. Genetics 176: 2371–2379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Pavlidis P, Jensen JD, Stephan W (2010) Searching for footprints of positive selection in whole-genome SNP data from nonequilibrium populations. Genetics 185: 907–922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Plagnol V, Wall JD (2006) Possible ancestral structure in human populations. PLoS Genet 2: 972–979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Slatkin M (2008) Linkage disequilibrium - understanding gthe evolutionary past and mapping the medical future. Nat Rev Genet 9: 477–485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Grossman SR, Shylakhter I, Karlsson EK, Byrne EH, Morales S, et al. (2010) A composite of multiple signals distinguishes causal variants in regions of positive selection. Science 327: 883–886. [DOI] [PubMed] [Google Scholar]
- 50. Sabeti PC, Reich DE, Higgins JM, Levine HZP, Richter DJ, et al. (2002) Detecting recent positive selection in the human genome from haplotype structure. Nature 419: 832–837. [DOI] [PubMed] [Google Scholar]
- 51. Voight BF, Kudravalli S, Wen X, Pritchard JK (2006) A map of recent positive selection in the human genome. PLoS Biol 4: e72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Ferrer-Admetlla A, Liang M, Korneliussen T, Nielsen R (2014) On detecting incomplete soft or hard selective sweeps using haplotype structure. Mol Biol Evol 31: 1059–65 DOI: 10.1093/molbev/msu077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Schierup MH, Hein J (2000) Consequences of recombination on traditional phylogenetic analysis. Genetics 156: 879–891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Úbeda F, Haig D (2004) Sex-specific meiotic drive and selection at an imprinted locus. Genetics 167: 2083–2095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Nielsen R, Bustamante CD, Clark AG, Glanowski S, Stackton TB, et al. (2005) A scan for positively selected genes in the genomes of humans and chimpanzees. PLoS Biol 3: 976–985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. da Fonseca RR, Kosiol C, Vinař T, Siepel A, Nielsen R (2010) Positive selection on apoptosis related genes. FEBS Lett 584: 469–476. [DOI] [PubMed] [Google Scholar]
- 57. Kosova G, Scott NM, Niederberger C, Prins GS, Ober C (2012) Genome-wide association study identifies candidate genes for male fertility traits in humans. Am J Hum Genet 90: 950–961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Seidel HS, Rockman MV, Kruglyak L (2008) Widespread gentic incompatibility in C. elegans maintained by balancing selection. Science 319: 589–594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Sellis D, Callahan BJ, Petrov DA, Messer PW (2012) Heterozygote advantage as a natural consequence of adaptation in diploids. Proc Natl Acad Sci USA 108: 20666–20671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Takahata N, Satta Y, Klein J (1995) Divergence time and population size in the lineage leading to modern humans. Theor Popul Biol 48: 198–221. [DOI] [PubMed] [Google Scholar]
- 61. Kumar S, Filipski A, Swama V, Walker A, Hedges SB (2005) Placing confidence limits on the molecular age of the human-chimpanzee divergence. Proc Natl Acad Sci USA 102: 18842–18847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Nachman MW, Crowell SL (2000) Estimate of the mutation rate per nucleotide in humans. Genetics 156: 297–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gillespie J (2004) Population genetics: a concise guide. Baltimore, MD: Johns Hopkins University Press, 2nd edition. [Google Scholar]
- 64. Pickrell JK, Coop G, Novembre J, Kudravalli S, Li JZ, et al. (2009) Signals of recent positive selection in a worldwide sample of human populations. Genome Res 19: 826–837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Hudson RR (2002) Generating samples under a Wright-Fisher neutral model. Bioinformatics 18: 337–338. [DOI] [PubMed] [Google Scholar]
- 66. Lohmueller KE, Bustamante CD, Clark AG (2009) Methods for human demographic inference using halptype patterns from genomewide single-nucleotide polymorphism data. Genetics 182: 217–231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Lohmueller KE, Bustamante CD, Clark AG (2011) Detectig directional selection in the presence of recent admixture in African-Americans. Genetics 187: 823–835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Marth GT, Czabarka E, Murvai J, Sherry ST (2004) The allele frequency spectrum in genome-wide human variation data reveals signals of differential demographic history in three large world populations. Genetics 166: 351–372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Drmanac R, Sparks AB, Callow MJ, Halpern AL, Burns NL, et al. (2009) Human genome sequencing using unchained base reads on self-assembling DNA nanoarrays. Science 327: 78–81. [DOI] [PubMed] [Google Scholar]
- 70. Pemberton TJ, Wang C, Li JZ, Rosenberg NA (2010) Inference of unexpected genetic relatedness among individuals in HapMap Phase III. Am J Hum Genet 87: 457–464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Wigginton JE, Cutler DJ, Abecasis GR (2005) A note on exact tests of Hardy-Weinberg equilibrium. Am J Hum Genet 76: 887–893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Hernandez RD, Williamson SH, Bustamante CD (2007) Context dependence, ancestral misidentification, and spurious signatures of natural selection. Mol Biol Evol 28: 1792–1800. [DOI] [PubMed] [Google Scholar]
- 73. Kong A, Thorleifsson G, Gudbjartsson DF, Masson G, Sigurdsson A, et al. (2010) Fine-scale recombination rate differences between sexes, populations and individuals. Nature 467: 1099–1103. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Performance of , , HKA, and Tajima's under the demographic models in Figure 2 with selection parameter and dominance parameter . Each row represents a different value. The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Mean difference in the number of polymorphic sites for a model with versus one with as a function of the distance from the site under balancing selection. Simulations were performed under the constant size divergence model in Figure 2A
with selection parameter , dominance parameter , and time of selection years ago. The mean difference in polymorphic sites is calculated for bins of size one kilobase and is plotted for 50 bins.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure 2 with selection parameter and dominance parameter . Each row represents a different value. The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure 2 with selection parameter and dominance parameter . The first panel is the divergence model in Figure 2A
. The second panel is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third panel is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of and under the constant size divergence model in Figure 2A
with no selected allele (neutrality). The first and second panels are scenarios in which we have erroneously over-estimated the recombination rate by two and one orders of magnitude, respectively (i.e., we respectively assumed recombination rates of and per base per generation when the simulations were performed using a rate of per base per generation). The third and fourth panels are scenarios in which we have erroneously under-estimated the recombination rate by one and two orders of magnitude, respectively (i.e., we respectively assumed recombination rates of and per base per generation when the simulations were performed using a rate of per base per generation). False positive rate is determined by neutral simulations under a model with recombination rate of per base per generation.
(PDF)
Performance of and under the constant size divergence model in Figure 2A
with selection parameter , dominance parameter or 1.5, and time of selection years ago. The first and second columns are scenarios in which we have erroneously over-estimated the recombination rate by two and one orders of magnitude, respectively (i.e., we respectively assumed recombination rates of and per base per generation when the simulations were performed using a rate of per base per generation). The third and fourth columns are scenarios in which we have erroneously under-estimated the recombination rate by one and two orders of magnitude, respectively (i.e., we respectively assumed recombination rates of and per base per generation when the simulations were performed using a rate of per base per generation). False positive rate is determined by neutral simulations under a model with recombination rate of per base per generation.
(PDF)
Demographic models used in simulations in which a selected allele arises prior to the split a pair of species. (A) Divergence model. Model parameters are a diploid effective population size , divergence time of the ingroup and outgroup species, and the time when the selected allele arises. (B) Divergence model with a recent bottleneck within the ingroup species. Additional model parameters are the diploid effective population size during the bottleneck, the time when the bottleneck began, and the time when the bottleneck ended. (C) Divergence model with recent population growth within the ingroup species. Additional model parameters are the current diploid effective population size after recent growth and the time when the growth occurred.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure S7 with selection parameter and dominance parameter . Each row represents a different value. The first column is the divergence model in Figure S7
A. The second column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure S7 with selection parameter and dominance parameter . Each row represents a different value. The first column is the divergence model in Figure S7
A. The second column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure 2 with selection parameter , dominance parameter , and time of selection . The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure 2 with selection parameter , dominance parameter , and time of selection . The first column is the divergence model in Figure 2A
. The second column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The third column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure 2 with selection parameter and dominance parameter . Each row represents a different value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure 2 with selection parameter and dominance parameter . Each row represents a different value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure S7 with selection parameter and dominance parameter . Each row represents a different value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size individuals. The first column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure S7 with selection parameter and dominance parameter . Each row represents a different value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size individuals. The first column is the divergence model in Figure S7
B with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure S7
C with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure 2 with selection parameter , and dominance parameter , and time of selection . Each row represents a different value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Performance of , , HKA, and Tajima's under the demographic models in Figure 2 with selection parameter , and dominance parameter , and time of selection . Each row represents a different value. The population sizes for these demographic histories have been scaled so that they produce the same number of segregating sites as a constant size population with diploid effective size individuals. The first column is the divergence model in Figure 2B
with a recent bottleneck within the ingroup species. The second column is the divergence model in Figure 2C
with recent population growth within the ingroup species.
(PDF)
Manhattan plot of genome-wide scans for balancing selection within the CEU population using the test statistic. From bottom to top, the horizontal dotted gray lines indicate the , , , and empirical cutoffs, respectively. The -axis is truncated at log composite likelihood ratio of zero.
(PDF)
Manhattan plot of genome-wide scans for balancing selection within the YRI population using the test statistic. From bottom to top, the horizontal dotted gray lines indicate the , , , and empirical cutoffs, respectively. The -axis is truncated at log composite likelihood ratio of zero.
(PDF)
Manhattan plot of genome-wide scans for balancing selection within the CEU population using the test statistic. From bottom to top, the horizontal dotted gray lines indicate the , , , and empirical cutoffs, respectively. The -axis is truncated at log composite likelihood ratio of zero.
(PDF)
Manhattan plot of genome-wide scans for balancing selection within the YRI population using the test statistic. From bottom to top, the horizontal dotted gray lines indicate the , , , and empirical cutoffs, respectively. The -axis is truncated at log composite likelihood ratio of zero.
(PDF)
Signals of balancing selection within the HLA region for the CEU (blue) and YRI (orange) populations using the test statistic. From bottom to top, the horizontal dotted gray lines indicate the , , , and empirical cutoffs, respectively.
(PDF)
Signal of balancing selection at the FANK1 gene for the CEU (blue) and YRI (orange) populations using the test statistic. From bottom to top, the horizontal dotted gray lines indicate the , , , and empirical cutoffs, respectively. SNPs (rsIDs) correspond to markers showing significant levels of transmission distortion within the Meyer et al. study.
(PDF)
Signal of balancing selection at the FANK1 gene for the CEU (blue) and YRI (orange) populations when removing either transitions or all transitions. SNPs (rsIDs) correspond to markers showing significant levels of transmission distortion within the Meyer et al. study.
(PDF)
Genealogy at the site of balancing selection.
(PDF)
Haplotype trees based on randomly sampling 18 haplotypes without replacement from a random simulation under the model in Figure S7
A. Trees were generated using UPGMA applied to a distance matrix of the proportion of nucleotide differences between each pair of haplotypes. The -kilobase (kb) window represents a region that is kb in length and is centered in the middle of the haplotype.
(PDF)
Top 100 signals in the CEU population using the test statistic.
(PDF)
Top 100 signals in the YRI population using the test statistic.
(PDF)
Top 100 signals in the CEU population using the test statistic.
(PDF)
Top 100 signals in the YRI population using the test statistic.
(PDF)
GO process analysis of top 100 signals, when compared to all signals, from CEU population using the test statistic.
(PDF)
GO process analysis of top 100 signals, when compared to all signals, from YRI population using the test statistic.
(PDF)
GO process analysis of top 100 signals, when compared to all signals, from CEU population using the test statistic.
(PDF)
GO process analysis of top 100 signals, when compared to all signals, from YRI population using the test statistic.
(PDF)
GO function analysis of top 100 signals, when compared to all signals, from CEU population using the test statistic.
(PDF)
GO function analysis of top 100 signals, when compared to all signals, from YRI population using the test statistic.
(PDF)
GO function analysis of top 100 signals, when compared to all signals, from CEU population using the test statistic.
(PDF)
GO component analysis of top 100 signals, when compared to all signals, from CEU population using the test statistic.
(PDF)
GO component analysis of top 100 signals, when compared to all signals, from YRI population using the test statistic.
(PDF)
GO component analysis of top 100 signals, when compared to all signals, from CEU population using the test statistic.
(PDF)
GO component analysis of top 100 signals, when compared to all signals, from YRI population using the test statistic.
(PDF)
GO process analysis of ranked signals from CEU population using the test statistic.
(PDF)
GO process analysis of ranked signals from YRI population using the test statistic.
(PDF)
GO process analysis of ranked signals from CEU population using the test statistic.
(PDF)
GO process analysis of ranked signals from YRI population using the test statistic.
(PDF)
GO function analysis of ranked signals from CEU population using the test statistic.
(PDF)
GO function analysis of ranked signals from YRI population using the test statistic.
(PDF)
GO function analysis of ranked signals from CEU population using the test statistic.
(PDF)
GO function analysis of ranked signals from YRI population using the test statistic.
(PDF)
GO component analysis of ranked signals from CEU population using the test statistic.
(PDF)
GO component analysis of ranked signals from YRI population using the test statistic.
(PDF)
GO component analysis of ranked signals from CEU population using the test statistic.
(PDF)
GO component analysis of ranked signals from YRI population using the test statistic.
(PDF)